끓는 물과 같다! 그래서, 불안정할 수 밖에 없고, 또한 그래서 불완전할 수 밖에 없다! (이 설명은 생기론, 물질론, 창발론에 대한 나의 절충적 대안이다!)
Life is thought to be ‘the boiling water,’ therefore it cannot help but being unstable and far from being complete! (This is my professional eclecticism on vitalism, materialism, and emergentism!)
(2021년 5월 28일 작성, 술 취한 김에… 증빙을 위해…)
아래는 2008년 12월 1일 기사입니다~
(원문: 여기를 클릭하세요~)
과학난제, 생명이란 무엇인가?장회익 교수, 물질현상과 생명현상 강연
인문학과 과학이 서로 협력, 미래를 만들어가는 인문강좌 행사가 최근 줄을 잇고 있다. 가장 주목을 받고 있는 행사는 한국학술진흥재단이 주최하는 ‘석학과 함께 하는 인문강좌’. 학문 간 경계를 넘어, 세상과 대화를 시도하려는 적극적인 노력이 엿보이고 있다. <사이언스타임즈>는 석학들이 진행하는 인문강좌를 연재한다.

우리는 대체로 지구상에 있는 여러 물리적 대상들 가운데 ‘살아 있는 것’과 ‘살아 있지 않은 것’을 구분하고, ‘살아 있음’을 특징짓는 성격을 지칭해 ‘생명’이라 부르고 있다. 또한 우리는 이러한 대상들이 ‘살아 있는 상태’에서 ‘살아 있지 않은 상태’로 전이되는 것을 ‘죽는다’고 말한다.
그러나 언뜻 보아 별 탈이 없어 보이는 이러한 생명 개념이 실제로는 적지 않은 문제점을 안고 있다.
29일 서울역사박물관에서 열린 석학과 함께하는 인문강좌에서 장회익 서울대 명예교수는 ‘물질현상과 생명현상‘이란 제하의 강연을 통해 “흥미롭게도 찰스 다윈(Charles Darwin의 할아버지였던 에라스므스 다윈(Erasmus Darwin)이 문제점을 잘 지적하고 있다”고 말했다.
새끼는 새로운 개체라고 할 수 없어
1794년 에라스므스 다윈은 새끼(offspring)를 새로운 동물이라고 부르는 데 대해 의문을 제기했다. 그리고 새끼는 새 동물이 아니라 어미의 한 가지(branch)이거나 돌출 부분(elongation)으로 보아야 한다고 주장했다.
왜냐하면 태 안에 들어 있는 동물은 어미의 기질을 일부 가지고 있는, 어미의 한 부분이었기 때문이다. 엄격히 말해 출생한 새끼는 새로운 개체라고 할 수 없다는 것인데, 이날 강연에서 장 교수는 “어미와 새끼를 언제부터 분리해야 하는지의 문제가 생명 단위와 관련해 훨씬 더 복잡한 문제를 제기하고 있다”고 말했다.
“토끼 안에 생명은 하나가 있는가?”
“아니면 세포의 수로 따져 약 천억 개의 세포가 있는가?”
“꺾어진 나뭇가지는 살아 있는 것인가?”
“아니면 죽어 있는 것인가?”

그렇다고 토끼로부터 분리된 세포들을 죽었다고 볼 수 없다고 말했다. 세포들은 비록 토끼로부터 분리됐다 하더라도 스스로 살아 있으며, 이를 통해 또 다른 복제가 가능하다. 살아있는지의 여부를 통해 우리가 판단하고 있는 ‘생명’의 존재 여부는 불확실하기 그지없다는 것이 장 교수의 설명이다.
그동안 많은 과학자들은 생명현상을 설명하기 위해 많은 노력을 기울여왔다. 그 중에서도 물리학자이면서 후에 생명연구에 본격적으로 뛰어들었던 우크라이나의 니콜라스 라세프스키(Nicholas Rashevsky)는 1950년대 생명의 본질을 이해하는 가장 중요한 공식을 소개한 중요한 인물이라고 장 교수는 말했다.
그는 “생명현상에 수학적 모형이 사용되는 물리학적 원리를 적용하는 것이 별로 유용하지 않으며, 유기체(organism)와 유기적 세계 전체(organic world as a whole)의 생물학적 일체성 (biological unity)을 나타내는 원리를 모색해야 한다”고 주장했는데, 그의 이 발언은 생명현상에 대한 강한 암시를 주고 있는 것이라고 장 교수는 평가했다.
반면 로젠(Rosen), 마투라나(Maturana), 바렐라(Barela) 등 라세프스키 이후의 많은 생명 연구는 거의 모두 실패했다고 말했다. 실패의 주된 원인은 생명현상을 설명하는 데 있어 ‘유기체’에 관련된 부분만을 설명하고 있을 뿐, ‘유기적 세계 전체’에 대한 연구를 간과해온 결과라는 것.
장 교수는 “생명의 진정한 모습은 서로 간에 긴밀한 연결망을 이루면서 그 안에서 ‘생명현상’이 진행되는 체계 전체를 하나의 실체로 파악했을 때 비로소 나타나는 것”이라며, 자신은 이 체계를 이전까지의 생명 개념과 구분, ‘온생명(global life)’이라고 불러오고 있다고 말했다.
생명현상, 온생명적인 접근이 필요해
장 교수는 ‘온생명’에 대해 “더 이상 분할하면 생명현상으로의 존립이 가능하지 않다는 점에서 생명이 갖추어야 할 최소의 단위임과 동시에 더 이상 외부로부터의 결정적인 지원이 없이도 생존을 해나갈 수 있다는 의미에서 생명이 지니게 되는 자족적 단위이기도 하다”고 설명했다.
만일 하나하나의 세포들은 ‘온생명’ 안에서 ‘온생명’의 나머지 부분과 적절한 관계를 유지할 때에 한해 살아 있다고 말할 수 있는 생명의 조건부적인 단위가 되며, 이를 ‘온생명’과 구분, ‘낱생명’이라고 부를 수 있다고 말했다.

최근 주목을 받고 있는 카프라(Capra) 역시 마투라나와 바렐라의 ‘자체생명성’을 생명의 정의로 채택하고 있는데, 많은 연구자들의 눈에 ‘낱생명’이 결코 자체생성적일 수 없다는 명백한 사실이 간과돼 왔다는 점은 쉽게 납득할 수 없는 일이라고 말했다.
장 교수는 자연계에 적용되는 열역학 제2법칙(물리학적으로 모든 열역학과정이 일어나는 방향이 정해져 있다는 법칙)에 조금만 관심을 돌려보더라도 자체생성성 개념은 원천적으로 ‘낱생명’에 대해서도 성립될 수 없는 이론이라고 지적했다.
장 교수는 생명현상을 설명하기 위해서는 낱생명적인 접근이 아닌 온생명적인 접근이 필요하다고 주장했다.
만일 생명이 출현하지 않은 우주의 여러 곳을 돌아다니며, 자연을 구성하는 모든 것의 존재 양상을 충분히 익힌 ‘우주인’이 있다고 가정했을 때 이 우주인이 지구상에서 일어나고 있는 생명현상을 어떻게 생각할 것인가.
물리학자 루드비히 볼츠만(Ludwig Boltzmann)은 1886년에 “생명체가 생존하기 위해 애쓰는 것은 원소들을 얻기 위해서가 아니다. 오히려 엔트로피를 얻기 위해서이며, 이것은 뜨거운 태양에서 차가운 지구로의 에너지 흐름을 통해 얻을 수 있다”고 말했다.
장 교수는 볼츠만(Boltzmann)이 생명현상에 대한 놀라운 해답을 던져주고 있다고 평가했다. 또한 볼츠만의 이론을 채용하고 있는 ‘생명이란 무엇인가?’의 저자인 슈뢰딩거(Schrödinger), 그리고 같은 제목의 책을 저술한 마굴리스(Margulis)와 세이건(Sagan)의 예를 들면서 이들을 통해 생명의 온생명적인 성격이 놀랍도록 선명하게 표현되고 있다고 말했다.
아래는 2010년 7월 13일 한 블로거의 글입니다~
(원문: 여기를 클릭하세요~)
생명의 정의
자연적인 ……
■ 생명의 정의 (A definition of life)
. 생명이란?
. 자기 복제 능력을 가지고
. 다윈적 진화를 하는 능력을 갖춘 화학 시스템이다.
→ 어떠한 화학 시스템이 생명을 특징 지우고 있는지?
→ 초기의 지구에서 원시 생명은 어떠한 에너지원을 이용했을까?
. 생명(生命)은 생물이 태어나서 죽기 전까지의 상태를 말한다.
. 생물학적 생명의 특성은?
. 성장한다. (↔ 바이러스는 성장하지 않는다.)
. 물질대사를 한다.
. 외부적으로나 내부적으로 움직인다. (↔ 불이 살아있다 ?)
. 자신과 닮은 개체를 생산해 내는 생식기능이 있다.
(↔ 노새는 생식 능력이 없다.)
. 외부 자극에 반응한다.
. 살아있는 생명체가 가지는 현상들은?
. 탄수화물, 지질, 핵산, 단백질과 같은 성분을 지니고 있다.
. 살아가기 위해 에너지와 물질을 모두 필요로 한다.
. 하나나 그 이상의 세포로 이루어져 있다.
. 항상성을 유지한다.
. 생명체의 종은 진화한다.
. 지구상의 생명체는 모두 탄소로 이루어진 유기체로 이루어져 있다.
. 어떤 사람은 이런 점이 모든 우주의 모든 생명체에도 해당한다고 보지만,
다른 한편에서는 이 현상을 ‘탄소 쇼비니즘’으로 부른다.
(참조 : From 위키백과, 須藤?2005 年度前期輪講レジュメ集 發表者:太田泰弘)
□ 생명의 정의
일반적인 생명의 정의로는 생리학적, 물질대사적, 생화학적, 유전적, 열역학적 정의
. 생리학적 정의; 섭식, 물질대사, 배설, 호흡, 이동, 성장, 생식, 외부자극에 대한 반응을 수행하는 계(系). 물질대사적 정의; 자신의 물질을 끊임없이 외부와 교환하지만 일반적인 특성을 잃지 않고 체제의 확실한 경계를 가지고 있는 물체. 생화학적 또는 분자생물학적 정의; 생명체는 핵산 분자에 생식 가능한 유전정보를 암호화하여 가지고 있고, 단백질성 촉매인 효소를 사용하여 물질대사의 화학반응 속도를 조절하는 계. 유전학적 정의; 자신들이 가지는 유기물질, 행동양식, 구조 등을 복제하는 존재. 열역학적 정의; 생명체는 개방된 계로 볼 수 있으며, 열, 빛, 물질 등 우주의 무질서를 통해 자신의 질서를 증가시키는 어떤 국소 부위로 정의
■ 생명의 화두
자료: Fm 인공생명(조.용현), 내용 중 일부 발췌
□ 근대과학과 환원론
환원론(reductionism)이란, 아무리 복잡한 것이라 하더라도 그것을 잘게 분해함으로써 기본적인 단순성에 도달할 수 있다고 하는 통찰에 기초 한 것이다. 이것의 가장 극명한 방법이 수학의 해석적(analytic) 방법이다.
직선은 단순하고 곡선은 복잡하다. 그러나 아무리 복잡한 곡선이라 하더라도 그것을 잘게 분해해서 확대해 보면 직선에 근사해 간다. 그래서 이 분할의 과정을 무한히 진행하면 곡률 0의 온전한 직선을 얻을 수 있다는 발상이다.
복잡한 곡선은 단순한 직선들의 집합에 지나지 않는다. 울통불통한 곡선으로 둘러쌓인 도형의 면적도 잘게 부수면 사각형들의 집합으로 환원될 것이고 이 사각형들의 면적을 합한다면 복잡한 도형의 면적도 쉽게 구할 수 있다.
환원주의의 전략을 무색하게 하는 악명높은 곡선 가운데 코흐곡선(koch curve)이라는 것이 있다. 이 코흐곡선은 다음과 같은 과정을 통해서 만들어 진다. 일단 임의의 길이를 취해서 그것을 3등분한다. 그 중간부분을 밑변으로 하는 새로운 삼각형을 그린다. 그리고 이것을 다시 3등분해서 앞서와 같은 방식으로 삼각형을 그린다. 이 과정을 되풀이 함으로써 코흐곡선을 얻을 수 있다.
이것은 고전적 의미의 곡선과 전혀 닮지 않았다. 고전적 곡선의 경우 작은 부분을 취하면 직선과 구분되지 않는다. 그러나 코흐곡선의 경우 아무리 잘게 잘라나가도 이 꾸불꾸불한 부분을 제거할 수 없다. 분할의 과정속에서 계속 전체의 복잡성이 재현되고 있을 뿐이다. 이러한 종류의 곡선은 깔끔한 방정식의 꼴로 나타낼 수 없기 때문에 “병리적 곡선”(pathological curve)이라고 불리어 졌는데 어째 그 이름에서 이치에 닿지 않는다는 의미인 “무리수”의 곤혹을 연상시킨다.
□ DNA와 환원론의 성공
수학이나 물리학에서는 이러한 현상이 나타나더라도 약간의 교정을 통해 전형으로 근사화 시킬 수 있기 때문에 예외적 현상으로 치부할 수 있다. 그러나 생물을 다룰 경우는 이러한 현상들은 드문 예외가 아니고 오히려 흔한 경우들이다. 그래서 생명의 현상은 환원주의 전략의 최후의 미개척지인 셈이고 그 완성을 확인하는 영역인 셈이다.
17세기 훅(R.Hooke)의 세포의 발견에서 시작된 환원주의적 전략은 20세기에 들어와서 왓슨과 크릭에 의한 DNA의 구조의 해명에 의해서 그 절정에 도달했다. 생명의 복잡성은 A,T,G,C라는 네 염기의 조합의 산물이라는 것이다. 이 중 세 염기의 조합으로 아미노산이 만들어지고 (가능한 조합의 수는 4×4×4=64종이지만 중복되는 것이 있어 자연계에는 20종이 있다) 다시 이 아미노산들의 조합으로 단백질이 만들어 진다. (아미노산에서처럼 염기수의 제한이 없으므로 원리적으로 무한수의 조합이 가능하다) 이 단백질들이 모여 세포를 구성하고 이 세포들이 모여 생명체의 몸을 구성한다.
이것은 생명일반이 보여주는 보편적 현상이면서 환원주의 전략을 무력하게 해 온 그래서 생기론의 강력한 논거가 되어온 생명의 자기재생산 현상을 설명해 주었다.
생명체가 생명체를 복사하는 것은 복사기가 서류를 복사하는 것과는 다르다. 그것은 자기자신을 복사하는 것이며 구태여 비교하자면 복사기가 복사기를 복사하는 것과 같다. 이것은 복사기가 자기 내부에 복사기 제조공장을 갖고 있어야 한다는 것을 의미한다. 그러나 이것은 아직 해결책이 아닌데 그렇게 해서 생산된 2세대 복사기는 다음세대의 복사기를 생산하지 못하는 불임의 복사기일 것이기 때문이다.
이것은 생명체의 복사와는 다르다. 생명체는 복사된 것이지만 자신도 복사를 수행할 수 있다. 그러므로 복사기가 생명체와 닮기 위해서는 복사기가 복사기와 함께 복사기 제조공장도 함께 복사해야 한다.
복사기의 복사의 연속성이 보장되기 위해서는 복사기 제조공장의 공장의 공장의…. 무한한 공장이 복사기 속에 내장되어 있지 않으면 안된다.
이것은 알속에 성체가 들어 있고, 알속의 성체속에 다시 알이 들어 있고, 다시 알속의 성체속의 알속에 성체가 들어 있는 등 무한히 축소되어가는 알과 성체의 계열을 인정해야 하는데 이것은 가능하지 않다. 이것이 소위 “정자미인”(精子微人, homunclus)의 역설이다.
이것은 환원주의(기계론)에 대한 생기론자들의 강력한 반대논거였지만 왓슨과 크릭의 DNA의 구조의 해명은 이 신비에 대한 환원주의적 설명을 가능하게 했다. 우선 생명체의 복사는 이중과정을 통해서 일어난다. 이것을 도식화하면 다음과 같다.
(1)
DNA → RNA → 단백질
(2)
(1)은 DNA가 DNA를 복사하는 것으로 복제(replication)라고 불리우는 과정이다. 이것은 말하자면 복사기 공장의 설계도를 복사하는 과정이다. (2)는 DNA에 제시된 설계도에 따라 복사기를 복사하는 과정으로 전사(transcription), 번역(translation)이라고 불리는 과정이다. 생명의 복사는 말하자면 복사기를 복사한 다음 그 복사기의 설계도를 함께 복사해서 넘겨주는 과정이라고 하겠다. 이렇게 해서 정자미인의 역설을 피할 수 있다.
이것은 생명의 자기재생산의 문제를 말끔히 해결해 주고 있는가? (1)과 (2)의 과정을 주도하는 것이 효소인데 이 효소 자체가 (1)과 (2)의 과정을 통해서 만들어져야 한다는 역설이 여전히 남아있다. 이러한 자기순환성은 생명현상이 가진 중요한 특징들 중의 하나인데 이것은 발생의 문제에서 전형적으로 드러난다.
수정란은 단세포에서 시작하여 2.4.8.16…으로 세포수가 급격히 증대되면서 분화가 시작된다. 그런데 여기서 생겨나는 의문은 똑같은 세포인데 어떻게 어떤 것은 근육세포가 되고 또 어떤 것은 혈세포가 되고, 골세포가 되고, 신경세포가 되는가 하는 것이다. 몸을 전체적으로 설계하는 프로그램세포가 따로 있는가? 환원주의 전략이 성립하기 위해서는 그래야 할 것 같다. 그러나 이러한 착상은 가망이 없다. 왜냐하면 이 프로그램세포와 일반세포를 구분시켜주는 것이 무엇인가 하는 의문이 생기고 이것을 해결하기 위해서는 다시 프로그램의 프로그램이 필요할 것이고 다시 프로그램의 프로그램의 프로그램이 요구되는 등 무한한 프로그램이 필요할 것이기 때문이다. 이것은 실제의 생명체가 사용하는 방법이 아니다.
어떤 세포가 특정의 어떤 세포가 되는 것은 그 세포의 유전자의 어느 부분이 읽혀지는가에 의존한다. 유전자의 읽는 방법을 결정하는 것은 직접적으로는 세포질인데 그 세포질은 세포의 경계조건에 따라서 결정된다. 즉 그 세포가 그 인근에 어떤 세포를 갖고 있는가 하는 경계조건이 그 세포의 분화를 결정한다.
이것은 순환적이다. 왜냐하면 a와 b가 인근해 있을 때 a는 b에 의해서 결정되지만 b는 다시 a에 의해서 결정되기 때문이다.
□ 전체와 부분의 순환성
환원주의 전략이 실패하는 곳에는 뚜렷한 공통점이 있다. 그것은 순환성이다. DNA는 효소를 규정하지만 DNA의 활성은 이 효소에 의해서 규정되고 있다. 세포a의 특성은 세포b의 특성에 의해 결정되지만 b의 특성은 다시 a에 의해서 규정된다. 일반적으로 순환논법은 무한퇴행으로 이끌며 한 논증의 실패의 논거가 된다. 그러나 자연의 복잡성은 하나같이 순환적 특성을 갖고 있다.
카우프만(S.Kauffman)은 세포의 분화에 어떤 프로그램이 있는 것도 아니며 그렇다고 요소 자체가 가진 어떤 생기에 의한 것도 아니라고 요소들간의 연결자체의 결과라는 것을 보여주기를 원했다. 그는 세포분화를 모형화한 실험을 설계했다.
그는 100개의 전구를 무작위적으로 연결시켰다. 그리고 한 전구의 켜지거나 꺼진 상태가 다른 전구의 켜지거나 꺼진 상태를 결정하는 규칙을 부여했다.
각 전구는 켜지거나 꺼지거나 둘 중의 한 상태를 취할 수 있기 때문에 전체적인 연결망의 가능한 상태는 2^100 또는 10^30이다. 여기서 특정한 한 상태가 다시 재현될 확률은 10 가운데 하나이기 때문에 불가능하다. 그러나 이런 예상을 뒤엎고 10번째에 극적인 변화가 일어났다. 11,12,13 번째 이후 14번째에서 다시 10번째의 상태로 복귀한 것이다. 그 뿐 아니라 이어서 11,12,13번째의 상태를 재현하면서 다시 10번째의 상태로 복귀하는 순환을 보여주었다. 연결방식을 달리하면 재현되는 패턴의 형태는 물론 달라진다. 그러나 주기순환을 보여준다는 점에서는 일치했다.그는 그 뒤 강력한 컴퓨터를 사용해서 요소의 수를 늘리면서 실험한 결과 상태순환의 수는 대략 요소들의 수의 제곱근이라는 것을 보여주었다. (앞서의 100개는 너무 작은 수여서 이 결과와 일치하지 않는다) 인간의 유전자들의 수는 약 10만개이므로 이것의 제곱근은 대략 316이다. 이것은 인간의 세포의 종류의 수 254개와 근사적으로 일치한다.이것은 무엇을 의미하는 것일까? 중요한 것은 요소가 아니고 그 요소들을 연결하는 연결방식이다. 이 연결에 의해서 요소와 요소, 요소와 전체간의 복잡한 상호되먹임 현상이 출현하고 이 과정에서 발생하는 것이 자기조직화이며 생명체는 이 자기조직화의 최고의 구현체이다.
□ 자기조직화
부분이 전체속에 들어있으면서 또 전체를 자신속에 가질 때 가능해지는 것이 ‘자기조직화’이다. 이것은 자기의 조건을 자기 스스로 부과한다는 점에서 외부의 강제에 의해서 만들어지는 질서와는 다르다.
레이저 이론을 탄생시킨 독일의 물리학자 하켄(H.Haken)은 자기조직 시스템의 간단한 예로서 레이저를 예로 들고 있다. 서로 마주 보는 거울이 있는 상자속에 에너지를 부여하면 상자 속 원자의 일부는 이전보다 에너지 준위(準位)가 높은 여기상태가 되어 광자를 방출한다. 광자는 여기 된 다른 원자에 충돌하고 거듭 광자가 방출된다. 처음에는 광자들의 파가 서로 간섭하여 복잡한 파형을 만들어 내지만 서로 위상이 달라 상쇄 되어버려 방출 되는 빛은 약하다. 그러나 점차 에너지의 강도를 높혀 가면 갑자기 어느 시점에서 광자들이 동일한 위상으로 정렬 되어 강력한 단일 진동수의 빛을 방출하는데 이것이 레이저 광선이다.
왜 이렇게 되는 것일까? 이것은 여기상태에 있는 많은 분자들의 내부 운동에 동조(同調) 현상이 생기기 때문이다. 이것이 하켄의 ‘예속원리'(slaving principle)인데 그는 이것이 실재의 자기조직 현상의 본질이라고 생각했다.
예컨대 두 대의 바이올린을 조율하여 한 대는 탁자위에 올려두고 다른 한 대를 가지고 음악을 연주한다고 생각해 보자. 연주하고 있는 바이올린 선과 똑 같은 선이 탁자위에 놓인 바이올린에서도 울린다는 것을 확인할 수있다. 다시 말해 G선을 연주하면 탁자 위에 놓인 바이올린의 G선도 같이 울린다. 왜 이런 일이 발생하는가? 첫째 바이올린에서 발생한 공기의 파동은 두번째 바이올린에 가서 부딪친다. 방출된 음과 똑같이 조율된 두번째 바이올린의 선은 우선적으로 그 파동의 에너지를 흡수할 수 있다. 그것은 그 파동의 진동수와 자신의 고유한 진동수가 같기 때문이다. 이러한 체계 속에서 전달되는 에너지는 당연히 가장 최적의 조건상태에서 전달 될 수 있다. 이러한 동조 현상을 공명(共鳴)이라고 한다.
이와 마찬가지로 여기 된 분자 속에서 일어나는 광자의 왕복운동도 일정한 주기를 갖고 있기 때문에 같은 주기로 변화하는 힘을 밖으로부터 부여하면 그 힘에 동조해서 진동하도록 되는 것이다. 광자들은 서로 닮아감으로서 동조적으로 진동한다. 이것은 요소들 간의 상호작용이다. 그러나 자기조직화가 이루어지기 위해서는 이것만으로는 부족한데 그 동조상태의 질서는 외부 요인에 의해 쉽게 파괴될 수 있기 때문이다. 그러나 일단 동조상태에 들어가면 그 체계는 외부요인에 저항하며 그것을 유지하는 힘을 갖고 있다. 그것은 왜일까?
또 다른 보기를 들어 설명해 보자. 추의 길이가 같은 구식 괘종시계가 여러 개 있다고 하자. 처음에는 추는 각기 제멋대로 움직인다. 시간이 지남에 따라 모든 추들이 마치 함께 묶여서 움직이듯이 일제히 같은 방향, 같은 속도로 움직이는 것을 볼 수 있을 것이다. (이것은 앞서 논의한 동조 현상과 같다) 그런데 그 가운데 하나의 추를 건드려 다르게 움직이게 해 보자. 얼마 안가서 또다시 다른 시게와 리듬을 맞추는 것을 보게 될 것이다. 이제 이것들은 질서의 교란에 대해 저항하고 계의 일체성을 유지하고자 한다. 구성 요소가 많을수록 계는 더 교란시키기 어려운데 제멋대로 움직이는 요소는 그 질서를 강요받는 듯이 보인다. 이것은 요소들의 운동에 의해서 전체 수준의 계가 생성되었지만 이제 역으로 그것에 작용하여 그것을 규정하고 있다. 이것이 하켄에 의하면 자기조직적 계가 출현하는 원리이다.
자기조직은 전체와 부분간의 상호 되먹임의 결과이다. 즉 처음에는 어떤 부분이 인접해 있는 다른 부분에 자신을 동조시킴으로 계의 일정한 특성을 만들고 이것이 다시 부분들에 작용함으로써 그 계의 특성을 강화시킨다. 이러한 되먹임은 자기가 자기를 만드는 촉매의 기능을 하고 있다는 점에서 ‘자기촉매성'(autocatalysis)이라고 불린다.
이 하켄의 자기 조직화의 원리는 프리고진으로 대표되는 브뤼셀학파(Brusselator)에 와서 더 넓은 범위에서 더 정교하게 다듬어졌다. 프리고진(I.Prigogine)에 의하면 평형에서 먼 혼돈은 자기조직의 가능성을 내포하고 있다. 그는 만일 열평형에서 멀리 떨어져 나올 수 있다면 많은 시스템들이 자발적으로 자신을 조직화 한다는 것을 증명해 주었다. 이렇게 해서 만들어진 구조들은 자기 조직화의 과정에서 계속적으로 엔트로피를 산출하고, 그에 따라 발생하는 엔트로피를 주변으로 퍼뜨린다고 해서 산일구조(散逸構造; dissipative structure)라고 불린다. 그 대표적인 것이 유명한 ‘벨로우소프-자보틴스키(Belousov-Zhabotinsky) 반응’이다.
시료들 중 하나의 농도가 임계점까지 증가되면 화학작용은 변환 되어 화학적 농도가 마치 화학시계처럼 규칙적으로 요동하기 시작한다.
수 십 억개의 분자들의 활동으로부터 유래되는 이러한 정도의 질서는 믿을 수 없는 것으로 보이며, 사실상 화학시계들이 관측되지 않았다면 아무도 그러한 과정이 가능하다고 믿으려고 하지 않을 것이다. 단번에 색을 바꾸기 위하여 분자들은 ‘교신’할 수 있는 방법을 지녀야만 한다. 계는 전체로서 행동해야만 한다. 우리는 화학에서 신경생리학에 이르기 까지 너무나 많은 분야에서 명백하게 중요한 교신이라고 하는 이 중요한 단어를 계속해서 접하게 될 것이다. 산일 구조들은 교신을 위한 가장 간단한 물리적 기구들 중의 하나를 도입하는 것이다.”
이 과정에서 자기와 같은 형태의 분자가 미리 존재하지 않는다면 그 분자가 만들어지지 않는다. 그와같은 화학물질은 자기자신의 촉매가 되는 것이다. 비유적으로 이야기 한다면 한쪽으로 ‘붉은’ 분자들이 집중되는 우연한 기회가 있으면, 이 ‘붉은’ 분자들은 더 많은 ‘붉은’ 분자들이 생성하도록 촉매역할을 하며 이렇게 생성된 2차적 ‘붉은’ 분자들은 또 새로운 붉은 분자들을 만들기 위한 촉매가 된다. 즉 반복 점진적으로 진행하는 것이다.
요컨대 요소와 요소 간, 요소와 전체 간의 자기되먹임의 메카니즘이 자기 조직화의 비밀인 것이다. 이러한 자기 조직화는 주변에서도 쉽게 관찰해 볼 수 있다.
비근한 예로 무리를 지어 날아 다니는 모기 떼의 경우를 보자. 모기 떼들은 분산도 확산도 하지 않으면서 그 형태를 아주 잘 유지한다. 개별적인 모기들의 운동의 무질서한 성격을 고찰해 볼 때 이것은 놀랄만한 점이다. 이것은 요소와 요소간의 상호작용(다른 모기떼로부터 떨어지지 않으려는 성향)과 집단과 요소간의 상호작용(집단적 운동은 개별 모기의 무질서한 운동을 방해한다)이 결부되어 만들어 지는 질서이다.무작위적인 움직임이 되먹임을 통해서 복잡한 질서에 이르는 보다 고차원적 과정은 흰개미의 집짓기에서 볼 수 있다. 흰개미들의 집짓기에는 아무런 설계도도, 그것을 감독할 중앙관료 제도도 없다. 처음에 개미들은 흙 덩어리를 물어 들고 이리저리 옮기면서 무질서하게 돌아다닌다. 이렇게 하면서 다른 개미들을 끌어 모으는 페르몬이라는 화학물질을 방사해서 흙 더미에 스며 들게 한다. 되는 대로 하다가 어느 곳에 흙이 더 많이 쌓이고, 이렇게 되면 그곳이 다른 개미들과 그들이 나르는 흙 덩이가 모이는 촛점이 된다. 그것은 더 많은 개미들을 끌어 들이고 개미의 행동은 이제 그 구조에 구속되게 된다.
인간사회에서 옷의 패션의 변화도 이러한 자기되먹임의 과정을 잘 보여준다. 패션계획 (비선형성, 불확정적) – 유행 – 강제성
□ 인다라망, 태극, 연결망
코흐곡선과 같은 프랙탈 도형들이 가지는 특징은 아무리 잘게 분할해 가도 그 속에 전체의 형태가 닮겨 있다는 것이다. 이것은 부분이 전체보다 크다는 의미에서 형식논리의 차원의 오류를 범하고 있다. 그러나 이러한 통찰들은 의외로 많다.
우선 라이프니쯔의 모나드(monad)의 개념이다. 라이프니쯔에 의하면 각각의 모나드의 표상속에는 일체의 다른 것과의 연관 즉 우주 자체가 함의되어 있다. 이러한 의미에서 하나하나의 모나드는 극도로 압축된 우주 즉 하나의 소우주(microcosmos)라고 할 수 있다.
법장은 유명한 금사자상(金獅子像)에서 이와 유사한 논의를 펴고 있다. “사자의 각 갈기 마다 금사자가 들어 있다. 또 그 갈기 안의 사자의 갈기 안에 또 사자가 들어 있다. 이런 식으로 계가 계를 포용하는데는 끝이 없으며 이것을 인다라 경계문(因陀羅境界門)이라 부른다” 또 화엄경에서는 “일체의 세계가 한 터럭만한 데 들어가고 한 터럭만한 데에 일체의 세계가 들어가며 일체 중생의 몸이 한 몸에 들어가고 한 몸이 일체중생의 몸에 들어가며 말할 수 없는 劫이 한 생각에 들어가고 한 생각에 말할 수 없는 겁이 들어가며 한 순간에 삼매(三昧)에 들어가 억겁이 일어나고 억겁이 들어가 한순간에 일어난다”고 하여 공간의 세계는 물론 시간의 세계까지 전체 속에 부분이 들어오는 한편 그 부분속에 전체가 들어오는 프랙탈적 측면을 강조하고 있다.
부분 속에 전체가 들어 있다는 이러한 논의들이 역설처럼 보이는 것은 우리가 은연중에 이것을 실체론적 관점에서 이해하고 있기 때문이다. 이것은 만물이 관계의 망 속에 얽혀 있다는 통찰의 비유적 표현으로 보아야 한다. 이 세계의 어느 부분도 고립되어 존재하는 것은 없다. 모든 것은 다른 것과의 관계 속에서만 존재한다. 그러므로 어떤 부분도 자신을 포함한 우주 전체와 연관되어 있고 그것을 반영하고 있다. 어떤 부분을 고립시켜 보면 그 부분속에 전체가 들어 있는 것처럼 보인다. 이것은 관계의 양상을 실체의 언어로 표현한 것이며 우주의 인다라망의 언어라는 거울에 비친 반영이다.
우주의 모든 것은 거울이면서 동시에 상이다.
그것은 만물을 반사하므로 거울이고 동시에 모든 것에 의해 반사되므로 상이다. 말하자면 하나의 사물은 다른 모든 사물과 관련되어 있기 때문에 그들 모두를 반영해 내며 어떤 특정사물은 다른 사물에 의존하고 있기 때문에 그것은 그 자체라기 보다도 다른 대상의 상이라고 볼 수 있다. 이것이 불교의 해인삼매(海印三昧)인데, 라이프니쯔의 모나드도 이와 다르게 보이지 않는다. (모나드가 창이 없다는 것은 거꾸로 뒤집어 보면 모나드 자체가 창이기 때문이라고 볼 수 있다)
부분과 전체의 역동적 상호관계는 주희의 이분수설에서도 볼 수 있다. 태극(太極)에서 이(理)와 기(氣)가 분화되어 나온다. 그런데 주희는 이 기속에 태극이 들어 있다고 한다. 이것은 태극과 기가 단순한 부분과 전체의 관계가 아니라는 것을 의미한다.
주희의 문제가 수정난에서 세포의 분화와 씨름한 카우프만의 문제와 크게 다르지 않다고 본다.
이것은 동질적인 것에서 이질적인 것의 분화의 문제이기 때문이다. 주희의 경우 개별화의 원리인 기속에 태극이라는 우주가 들어오고 있다. 이것을 카우프만의 표현방식으로 말하면 부분과 전체의 역동적인 관계 맺음이다. 반면 카우프만을 주희의 언어로 표현하면 전체의 네트워크가 태극이고 그것은 분화된 개별자 속에 함의되어 들어있다.
이러한 의미에서 인다라망(불교), 태극(주희), 관계망(카우프만)은 바로 생명의 본질을 표현한 것이고 전체가 부분 속에 내재해 들어오는 자기조직의 역동적 관계 속에서 사물을 파악하고 있다는 점에서 동일하다고 하겠다. 이러한 착상들은 실체를 기본 개념으로 하고 상호 고립적인 물질을 대상의 범형으로 파악한 서양과학의 환원론적인 전통과는 구분된다. 이것은 아주 오래된 통찰이지만 최근 비평형 열역학, 혼돈이론, 인공생명 등을 통해서 과학의 주제로서 새삼스럽게 주목받으며 재발견되고 있다.
■ 구멍 주름 생명
자료 Fm : 조.용현 (내용 중 일부 발췌)
……
생명은 그 본성상 “관계자”이다.
그것은 스스로 관계 맺어 자기조직화 하려고 하는 경향을 가진다.
거기에는 바깥으로부터의 설계자가 없다. 요소와 전체의 연관은 내적인 필연성으로 이루어진다. 그것은 바깥으로 자기를 열어놓고 있고 그래서 표면은 면적을 최대화하기 위해 울퉁불퉁한 구조를 취하고 있다.
기계에서 “관계”는 죽음의 방식이지만 생명에서 “관계”는 삶의 방식이다.
생명체들의 외양상의 매끄러운 모습에 현혹되어서는 안 된다.
구멍과 주름이 몸의 곳곳에 은폐되어 있다.
그것이 허파이고 핏줄이고 장이다.
그것은 피부의 확장이다. (이것들은 피부와 동일한 상피세포로 되어 있다.)
박테리아는 이렇다 할만한 내부 기관이 없다.
그것은 호흡계도, 순환계도, 소화계도 없다.
그것은 피부로 호흡하고 피부로 소화한다.
말하자면 피부로 “듣고”, 피부로 “냄새 맡고”, 피부로 “본다”
박테리아에게는 왜 이것이 가능한가?
그 이유는 단순한데 사이즈가 작다는 데서 온다.
사이즈가 작으면 부피에 대한 표면적의 비가 커진다.
박테리아에게는 내부 장기가 결여되어 있는 것이 아니라 그것이 필요 없는 것이다. 사이즈가 커지면 표면적이 줄어들어 불가피하게 복잡한 기관들을 들여 놓아야 한다. 지렁이는 순환계는 있지만 호흡계는 없다.
그것은 피부로 호흡한다. 더 커지면 순환계와 호흡계라는 내부 장기가 필요하다.
이것은 순환계도, 호흡계도, 소화계도 모두 피부의 확장이라는 것을 의미한다.
큰 동물에게서 이제 바깥의 피부는 각질의 보호막으로 남고
보고, 듣고, 먹고, 배설하던 그 민감한 진짜 피부는 안으로 욱여 들어가서 감각기관이 되고 소화기관이 되고 배설기관이 되었다.
그 두터운 각질 속에 주름과 구멍으로 된 생명의 원형은 의연히 보존되어 있다.
……
□ 구멍, 주름, 프랙탈
구멍과 주름은 표면적을 최대화하는 방법, 말하자면 관계를 극대화하는 방법이다.
이것을 이해하기 위한 도식으로 멩거 스폰지라는 프랙탈 도형을 살펴보자.
이것은 입방체를 3등분하면 27개의 작은 입방체가 만들어질 것이다.
그 가운데 중간부분의 작은 입방체를 버린다.
다음 다시 남아있는 20개의 작은 입방체를 각각 3등분해서
그 가운데 부분들을 버린다.
이 과정을 무한히 반복함으로써 만들어지는 것이 멩거 스폰지이다.
이 과정을 무한히 한다면 이 멩거스폰지는
유한한 크기에도 불구하고 그 면적은 무한대가 될 것이다.
반면 단계가 진행됨에 따라
부피는 지속적으로 줄어들어 종국에는 0이 되어 버린다.
크기가 없으면서 무한대의 표면적을 갖는 대상!
물론 실제 생명체도 무한한 표면적을 갖지 않으며
또 아무리 작다 하더라도 크기가 없는 것은 없다.
그러나 이것은 생명체의 구멍과 주름에 대한 근사한 모형으로 보인다.
우리 몸의 폐는 텅 빈 공기주머니가 아니다.
큰 가지가 점점 작은 가지로 갈라지면서
폐포에서 종결되는 수지상 구조로 되어 있다.
산소는 폐의 세포벽과 접촉해야만 적혈구에 의해서 흡수될 수 있기 때문에
허용되는 최대 부피 내에 가능한 한 넓은 표면을 가지는 것이 유리하다.
프랙탈 구조는 이 문제에 대한 해결책이다.
똑같은 원리가 우리 몸의
순환계에도, 소화관에도, 신경계에도 적용되고 있다.
똑같은 요구 때문에 우리의 대뇌피질도 주름져 있고,
우리의 손바닥도 주름진 손금으로 되어있다.
……
□ 가이아의 주름
원자는 사물에 대한 극단적 이상화이다.
엄밀한 의미에서 원자적 사물은 존재하지 않는다.
정도의 차이는 있지만 생명 아닌 모든 사물도 프랙탈적 구조를 갖는다.
이 세계에 관계 지워져 있지 않는 것은 아무것도 없기 때문이다.
바다의 파도는 그것이 부딪히는 땅과 관계하여 독특한 해변의 프랙탈구조를 만들고 모래는 바람과 관계 맺어 모래사막의 아름다운 무늬를 형성한다.
강의 물줄기는 땅과 대화해서 햇빛과 식물의 대화에서 생겨나는 것과 유사한 수지상의 강모양을 만든다. 그러나 엽맥에서 보이는 것과 같은 이중교차가 없다.
이것은 생명의 관계 맺음의 정도가 훨씬 더 긴밀하다는 것을 보여준다.
우리가 살아있는 만큼 뭍 생명을 품고 있는 이 지구도 살아있다.
NASA로부터 화성에서의 생명체의 가능성에 대한 조사를 의뢰 받았던
제임스 러브록(J.Lovelock)은 이런 의미에서 지구를 “가이아”(Gaia) – 희랍신화의 대지의 여신 – 라고 부를 것을 제안했다.
……
생명이 살기에 적합한 “여건으로서의 지구”가 아니고 생명이 만들어낸 것이 바로 지구다.
우리는 단순히 그 환경 속에 살고 있지 않다. 우리가 바로 그것이다. 이것이 러브록이 내린 결론이었다.
가이아의 주름은 무엇보다도 수림이다. 특히 열대 우림이다.
이것은 폭 3천마일로 가이아를 둘러싸고 있는 전체 육지면적의 14%에 해당하는 푸른 띠이다. 나무 자체가 태양의 주름이다
……
□ 가이아의 구멍
나무가 가이아의 주름이라면 갯벌은 가이아의 구멍이다.
갯벌은 미세한 구멍이 송송 뚫려있는 다공질의 점토들로 되어 있다.
그 넓은 표면적으로 해서 산소와 영양의 공급이 원활하고 이것이 많은 생명체들의 삶의 터전이 되고 있다.
갯벌과 같은 습지 1m2에서 벌어지는 광합성작용의 생산량은 연간 1만-2만5천 칼로리로 사막의 50배, 산림의 20배, 먼바다의 10배에 해당한다. 그리고 바다 생물들의 90%가 개펄에서 산란하고 그곳에서 생을 시작한다. 그야말로 가이아의 자궁이라고 할 만 하다.
열대 우림이 생명체들의 서식환경이라기 보다 그 자체 초유기체라고 보아야 하듯이 이 갯벌도 그 자체 살아있는 초유기체이다. 생명체가 없으면 열대 우림이 그 특성을 상실하듯이 갯벌도 생명체가 없으면 갯벌로서의 특성을 상실한다. 문자 그대로 그것은 죽음을 맞이한다.
……
□ 작은 가이아
나는 생명체를 “작은 가이아”라고 부르기를 좋아한다.
이 용어에는 생명에 대한 우주론적인 울림이 있다.
생명체들은 그 어느 것도 그 자체로서 하나의 우주이라는 함축이 그 언어 속에 들어 있다.
이것은 새로운 생각이라기 보다 우리 선조들이 가졌던 생명에 대한 관점이었다. 그러나 기계론의 위세 앞에서 우리는 여기에 대한 느낌을 잃어버렸다.
……
생명의 기본디자인으로서 황금분할의 의미와 기능
소라와 같은 연체동물(mollusc)의 패각과 식물의 잎차례(phyllotaxis)
……
기계론의 해독을 씻어내고 자연의 神性을 되찾게 해주어야 한다.
뭍 생명들 속에 들어있는 가이아를 볼 수 있는 감수성을 길러야 한다.
우리가 먹고 있는 것은 단순한 단백질의 덩어리가 아니고 우리를 길러준 가이아이고, 우주이고, 하느님이고, 부처의 몸이다.
우리의 삶은 이 가이아 또는 부처의 몸 보시에 신세지고 있는 것이며 그것은 우리가 미래에 되돌려주어야 할 업보이다.
길가에 피어있는 이름없는 들꽃 속에도 한 우주가 깃들어 있으며
그것을 함부로 꺾지 마라. 우리의 무심한 행동이 한 우주를 붕괴시키고 있다.
나는 영국의 시인 윌리암 블레이크(W.Blake)의 높은 감수성을 좋아한다.
그는 “순수의 전조”(Auguries of Innocence) 라는 시에서 다음과 같이 노래했다.
한 알갱이의 모래 속에서 세계를 보며 한 송이 들꽃 속에서 우주를 본다. 그대 손바닥 안에 무한을 쥐고 한 순간 속에 영원을 담아라.
모래나 들꽃 속에 우주가 들어있음을
이 시인은 높은 감수성으로 느낀다.
……
(비록 우리의 감수성이 그만 못하더라도)
소라의 貝殼에서라면 그 느낌을 가져올 수 있다.
소라는 그리이스 인들이 신의 비(divine proportion)라고 부른
그 황금비가 3차원상에 아름답게 각인되어 있는 우주이다.
소라뿐이겠는가? 피뿔고동, 비단고동(각시고동), 바지락, 백합에 각인 된
아름다운 등각나선의 곡선을 찬찬히 따라가 보라.
무엇인가 아련히 들려오는 생명의 노래를 들을 수 있을 것이다.
우리가 둘이 아님을 ……
“입장 바꿔 생각해봐”라는 말이 있다.
사자성어로 易地思之.
……
역지사지 참 좋은 말이다.
길을 걸어가며 흙의 입장을 생각해보고,
나무를 볼 때 나무의 입장을 생각해보며,
날아가는 새의 입장을 생각해보는 것 얼마나 좋은 행동인가?
허나 내가 간과한 것은 나무의 입장을 생각할 때 나무의 입장이 되어 나무의 가치관으로 보지 않았던 것이다.
크기와 형태에 따라 세계관이 다르고 사는 방법도 다르며 시간도 다르다.
내가 나무의 입장을 생각하면 뭘 하나?
결국 나의 눈으로 나무를 보는 것 밖에 안 된다.
나는 사물들의 입장만 생각했지 진정으로 그 사물이 되어보지 못했던 것이다.
……
■ 세포 자동자, 생명
자료: Fm 인공생명(조.용현), 내용 중 일부 발췌
□ 세포자동자란 무엇인가?
“세포자동자”(cellular automaton)1940년대 폰노이만(von Neumann)은 생명을 ‘합성’하는데 관심을 가졌다. 이 계획을 위한 중요한 단계로서 인공적인 자기재생산계를 연구했다. 그의 처음의 개념적 디자인은 기계적인 것이었다. 로봇이 호수위를 떠다니면서 거기서 에너지와 원료를 추출해서 자기자신의 복사본을 만드는 것이다. 폰노이만이 실제 그러한 것이 현실화되리라고 생각한 것 같지는 않다. 그의 진정한 목적은 ‘자기재생산’을 이해하는 것이었다.울람(S.Ulam)은 맨허턴 계획에 사용되던 컴퓨터를 사용해서 기하학적 패턴을 재귀적 방식으로 재생산할 수 있는 프로그램을 만들었다. 이것은 재생산의 참된 특징은 물리적 로봇의 3차원세계 보다 추상적인 기하학적 공간에서 더 쉽게 연구될 수 있다는 것을 시사한다. 이 시사가 폰노이만에게 중요한 개념적 돌파구를 열어주었다. ‘세포자동자 이론’이 출현한 것이다. 세포자동자를 규정하는 것은 크게 다음 3가지이다.
. 세포당 상태의 수 . 세포의 이웃 . 규칙
□ 세포자동자와 혼돈의 가장자리
……
주기적 질서에서 혼돈으로 가는데는 두가지 경로가 있음을 알 수 있다.
하나는 주기적 거동에서 혼돈적 거동으로 곧장 이행해 가는 것으로 이것이 ‘1차상전이'(first-order phase transition)이다. 다른 하나는 주기적 질서에서 혼돈으로 가지않고 그 둘사이의 경계점을 통과해 가는 경우이다. 이것이 ‘2차 상전이'(second-order phase transition)인데 랭턴은 여기가 바로 ‘생명의 자리’라고 보았다.
물리학에서의 상전이 현상과 연관지어 보면 그 의미를 잘 알 수있다. 우리가 주로 잘 알고 있는 것은 일차 상전이 현상이다. 예를 들어 얼음의 온도를 상온으로 올리면 얼음은 한꺼번에 물로 변한다. 이 때 일어나는 현상을 분자차원에서 보면 분자들이 질서와 혼돈 중 어느 하나를 선택하도록 강요되고 있는 것이다. 전이온도 보다 낮은 온도에서는 분자들이 결정구조의 질서(얼음)를 선택할 수 있을 만큼의 느린 속도로 진동하고 있다. 반대로 전이온도 보다 높은 온도에서는 분자들이 너무 심하게 진동하기 때문에 분자결합이 끊어지는 속도가 재결합하는 속도 보다 빨라져서 혼돈(물)을 선택하지 않을 수 없다.
전이온도 보다 높은 온도에서는 대부분의 분자들이 완전히 혼돈스러운 상태로 서로서로를 넘어 뒹굴어 다닌다. 그러나 뒹굴어 다니는 것들 중에는 무수히 많은 아주 작은 섬들이 있는데, 이 섬들은 질서있는 격자상태의 고체이며 그 가장자리에서 분자들은 끊임없이 용해되었다가 재결정화 된다. 이 섬들은 분자크기의 레벨에서 보아도 그렇게 크지 않으며 오래 지속되지 않는다. 그러므로 그 시스템은 아직 거의 혼돈상태이다.
그러나 온도를 낮추어가면 섬들이 점점 커지기 시작하고 그에 상응해서 지속 기간이 점점 길어진다. 혼돈과 질서간의 균형이 변하기 시작한 것이다. 온도를 전이온도 보다 더 내리면 이제 상황은 역전된다. 유체의 바다 속에 점점이 박힌 고체의 섬에서 고체의 대륙위에 점점이 산재해 있는 호수들로 변한다.
둘 사이의 전이점이 2차상전이가 일어나는 점이다. 질서있는 구조들이 혼돈스러운 유체의 양과 똑같은 양만큼 물질을 채우게 된다. 질서와 혼돈이 혼재되어 있으며 비유적으로 말하자면 각 분자들이 조직의 일원으로 남을 것인지, 외롭지만 자유로운 개인으로 남을 것인지를 결정하지 못하고 갈등상태에 빠져 있는 것과 흡사하다. 이 갈등은 상호타협-전체속의 부분, 부분속의 전체-를 낳게 되는데 이것이 바로 복잡성의 영역이다.
물리학에서의 상전이 현상에 대한 기술이 세포자동자에 대한 기술과 너무나 흡사해서 흥미롭다. 세포자동자의 λ는 물리학에서의 온도와 같은 역할을 하는 것인지 모른다. 그러나 더 흥미로운 것은 랭턴의 논의가 생명이 왜 물과 연관되어 있는지에 대한 중요한 시사를 던져주고 있다는 것이다.
물질은 고체, 액체, 기체라는 3가지 위상을 가진다. 그러나 액체에서 기체로 또 그역으로의 전이는 위상전이 없이 가능하기 때문에 액체와 기체는 더 일반적인 단계인 ‘유체'(fluid)라고 볼 수 있다. 고체와 유체는 위상전이의 산물이기 때문에 현상태의 현격한 차이에도 불구하고 이 둘은 동일한 실체의 다른 相에 지나지 않는다. 그러므로 고체와 유체의 차이는 그 구성물질에 있는 것이 아니라 그것의 동력학적 거동방식의 차이에 있는 것이다. 앞서의 지형도를 따르면 고체는 고정점과 주기의 영역에 있는 것이고 유체는 혼돈의 영역에 있는 것이다. 그리고 물(액체)은 유체로서의 기본위상을 가지면서도 고체와 유체의 경계선상에 위치하고 있다. 이 경계는 질서 속에 얼어 붙지 않으면서도 혼돈 속에 빨려들어 가지도 않는 그 미묘한 경계영역이다.
이곳이 바로 생명의 자리인데 랭턴은 이곳을 ‘혼돈의 가장자리'(edge of chaos)라고 불렀다.
□ 세포자동자와 생명의 의미
이것은 어떻게 생각하면 컴퓨터 게임에 지나지 않는 콘웨이의 라이프게임이 왜 생명적 거동을 보여주는지를 설명해 준다. 물질의 상을 기본적으로 동력학적 거동이라고 본다면 중요한 것은 그 거동이지 그것을 이루고 있는 소재가 아니다. 거동을 만들어내는 것이 무엇이든 -그것이 컴퓨터의 소프트웨어든 생물의 웨트웨어든- 그 거동이 동일한 위상-복잡성-속에 있을 때 동일한 양상-생명현상-을 보일 것이다.
“고체, 액체, 그리고 기체의 동력학적 계에서 거동의 보편적 집합으로서 일반화는 컴퓨터의 ‘하드웨어’에 의해서 구현되는 각종 거동들을 이해하는데 중요한 귀결들을 보여준다. 컴퓨터의 하드웨어는 생물의 ‘웨트웨어'(wetware)와는 근본적으로 다른 ‘소재’이며 이 근본적 차이 때문에 컴퓨터의 하드웨어는 생명, 지성과 같은 웨트웨어가 보여주는 한층 더 복잡한 거동을 성취할 수 없다고 일반적으로 생각되고 있다. 그러나 고체형, 액체형, 기체형이 물질 그 자체의 성질이라기 보다 물질의 조직화의 차이라는 것을 정확히 이해한다면 하드웨어를 웨트웨어로 바꾸는 것은 단지 조직화의 문제이며 결과적으로 웨트웨어가 성취한 모든 것을 하드웨어도 성취할 수 있다고 보아야 할 것이다.
부가해서 이러한 통찰은 위상전이의 근방에서 작동하는 물질계는 컴퓨터처럼 거동할 수 있다는 것을 함축한다. 이것은 웨트웨어에 의해서 성취될 수 있는 각종 거동들을 이해 할 수 있는 중요한 귀결들을 갖고 있다. ‘물’은 동력학적 용어로 말하면 질서와 혼돈 사이를 가로지르는 혼돈의 가장자리이고, 콘웨이의 라이프게임(23/3의 규칙)은 웨트웨어의 용어로 말하면 s=2, n=9가 만드는 광대한 규칙공간(대부분은 고착된 질서나 혼돈의 땅)속에 드물게 분포해 있는 ‘물’의 영역이다.
정보동력학에서의 ‘복잡성’은 생물학에서의 ‘생명’의 다른 이름이다.
우리는 정보동력학이 임계위상전이 근방의 물리계에서 자발적으로 출현할 수 있다는 것을 이제 알게 되었다. 이 논문의 결과는 국지적 또는 전역적 조건이 임계위상전이의 영역에 어떤 매질-그것은 물일지도 모르고 또 다른 것일지도 모른다-을 가져왔을 때 생명을 발생시킨 정보동력학이 나타났다는 것을 시사하고 있다. 우리가 세포자동자에서 임계위상전이의 부근에서의 동력학이 준안정적 구조의 복잡동력학을 조성시킨다는 것을 보았는데 이 준안정적 구조의 복잡동력학은 바로 생명을 특징지우는 바로 그것과 같은 종류의 것이다. 자연은 임계위상전이의 부근에 오랫동안 이 매질을 유지시켜줄 만큼 자비롭지는 않다. 이것은 초기의 정보 동력학이 전역적 조건이 위상전이영역을 떠났음에도 불구하고 국지적으로 위상전이의 근방에 머물 수 있도록 몇가지 매개변수들을 통제하고 있었음에 틀림없다는 추론을 가능하게 한다. 물론 그러한 계를 중요한 전이점으로부터 멀리 밀어내는 많은 매개변수들이 있었음에 틀림없고 그 종류도 아주 다양해서 이것들이 초기의 정보처리계의 대부분을 파괴시켰을 것이다. 진화란 중요한 위상전이에 대한 계의 관계에 영향을 주는 “매개변수”들에 대한 통제범위를 확장시켜가는 과정이라고 볼 수 있다.
살아있는 계란 동력학적으로 말한다면 ‘끌개’를 피할 수 있는 계라고 말할 수 있다. 주기영역은 한계순환이나 고정점 끌개에 의해서 규정되는데 대해 혼돈의 영역은 전형적으로 아주 많은 차원으로 된 이상한 끌개에 의해서 규정된다. 살아있는 계는 이 양 극단을 피해야 하고 그래서 지나친 질서와 지나친 무질서 사이의 교묘한 운행을 배우는 것이 필요하다.
이것은 연장된 표류기, 다른 말로 ‘임계’ 전이 근방에 자신을 머물게 하는 것을 배움으로써 가능하다. 일단 그러한 계가 임계전이 근방에서 출현하면 진화는 임계점 근방의 동력학의 본질인 정보처리능력을 획득하게 되고, 열려 있는 표류기에 자신을 유지할 수 있는 능력을 더욱 확장하기위해 그 능력을 이용해 왔던 것으로 보인다.”(C.Langton,”Artificial Life”in the philosophy of Artificial Life, p.39)
생명의 본질은 그 동력학적 거동에 있지 그 소재에 있는 것이 아니라는 랭턴의 주장이 타당하다면 우리는 생명연구에 있어서 지금까지의 어쩔 수 없었던 한계-지구상의 생명이라는 특수한 사례를 통한 생명의 보편적 속성에 대한 탐구라는 불가능한 과업-를 극복할 수 있는 새로운 돌파구를 찾은 셈이다. 컴퓨터 내에 복잡성을 구현해 봄으로써 생명의 본질에 대해 우리는 많은 것을 배울 수 있을 것이다. 세포자동자가 바로 그 한 예이다.
■ 카우프만의 “신의 마음”
Stuart Kauffman, At Home in the Universe : Oxford Univ.Press,1996
과학철학, 1999 (조.용현), 일부내용 발췌
물질적 우주에서 생명이란 도대체 어떻게 가능한가? 자신이 자신에 대해서 질문을 던지는 이 의식이라는 것이 어떻게 가능한가? 도대체 적어도 무엇이 “있다”는 것이 어떻게 가능한가?
요즈음 철학은 더 이상 이러한 질문에 흥미를 갖지 않는다. 고대 자연철학 이후 철학적 정신을 사로잡아왔던 이러한 주제들은 오늘날에 와서 이미 빛 바랜 유물처럼 되어 버렸고 영화와 같은 “일상적 잡담에서의 의미 찾기”가 이제 철학의 본령인양 되어 가고 있다.
□ 스튜어트 카우프만(S.Kauffman)의 『우주는 우리의 집』(at home in the universe)
그는 이 책에서 우연을 넘어서는 어떤 질서-그의 용어로 賦存秩序(order for free)-가 있음을 아주 설득력 있게 보여주고 있다. 이 책은 생명의 기원에서 시작해서 수정난의 형태형성에 이르기 까지, 캄브리아기의 대 번성에서 기술혁명에 이르기 까지 여러 다양한 주제들의 근저에 깔려있는 질서를 보여주고자 하고 있다. 그러나 무엇보다도 생명에 대한 반기계론적 대안으로서의 생기론을 대체 할 수 있는 강력한 대안이 될 수 있지 않을까 하는 생각을 가졌다. 필자는 생기론의 생명에 대한 직관이 틀렸다고 생각하지 않는다. 문제는 그 설명방식,즉 “生氣”(elan vital)라는 개념의 그 공허 한 동어반복에 있다.
“생명의 기원에 관한 나의 이론은 철저한 전체론(holism)에 입각하고 있다. 그러나 이 이론은 신비적인데서 나온 것이 아니고 수학적 필연성에서 도출된 것이다… 생명은 조금씩, 조금씩 만들어진 것이 아니고 전체로서 출현했으며 또 그렇게 유지되고 있다.”
생기론자도 아니고 더더구나 창조론자도 아니면서 어떻게 이런 주장을 할 수 있는가? 바로 여기에 이 책의 매력이 있다.
□ 유전자 연결망
카우프만은 특이한 경력을 가진 생물학자이다. 그는 다터마스 대학에서 철학을 공부한 뒤 옥스퍼드대학으로 유학해서 철학과 심리학을 전공했다. 그는 다시 의학으로 진로를 바꾸어 샌프란시스코 의대로 진학했다. 농담인지 모르겠지만 진로를 바꾸면서 다음과 같은 엉터리 삼단논법을 전개했다고 한다. “위대한 철학자가 되기 위해서는 임마누엘 칸트만큼 똑똑해야 한다. 나는 칸트만큼 똑똑하지 못하다. 그러므로 나는 의사가 되겠다.”
그는 의과대학에서 생물학을 공부하면서 수정난에서의 형태발생(morphogenesis)의 문제에 깊이 매료되게 된다. 어떻게 무규정적인 동질적 胚 세포에서 근육세포, 신경세포, 혈액세포 등이 분화되어 나올 수 있는가? 인간을 구성하는 10만종의 유전자가 어떻게 254개의 상이한 세포들을 발현하는가?
사실 이 문제는 19세기 바이스만(A.Weisman)으로 거슬러 올라가는 오랜 문제이며 기계론과 생기론의 전투의 현장이기도 했다. 모든 세포는 수정난에서 시작한다. 그것은 분화를 시작하면서 특정세포로 형성되어 가지만 특정세포가 다시 분화이전으로 되돌아가는 경우는 없다. 그는 발생이 한 방향으로만 진행되는 이유는 유전정보가 세포분화에 따라 줄어들기 때문이라고 생각했다. 그래서 수정난에는 완전한 개체를 생산하는데 필요한 모든 정보가 들어있지만 분열할 때 마다 정보가 반으로 줄어들게 된다. 이것은 아주 그럴듯한 설명이었다..
바이스만의 추론이 맞다면 수정난을 2세포 단계에서 한쪽을 파괴시키면 나머지 반쪽에서 반쪽자리 개체가 생겨나야 할 것이다. 한스 드리쉬(Hans Dreisch)가 성게의 알(카우프만은 개구리의 알이라고 말하고 있는데 이것은 그의 착각인 것 같다.)을 가지고 이 실험을 해본 결과 바이스만의 예측과는 달리 온전한 성게의 배로 발달했다. 어떻게 반쪽이 전체의 정보를 가질 수 있는가? 이것은 기계라면 절대로 보일 수 없는 현상인데 부분 속에 전체가 구유되어 있기 때문이다. 드리쉬는 발생을 유도하는 비물질적인 일종의 힘이 있다고 보고 이것을 아리스토텔레스의 용어를 빌려 “엔텔레키”(entelechy)라고 불렀다.
1960년대 들어와서 쟈콥(F.Jacob)과 모노(J.Monod)가 전개한 오페론(operon) 가설은 엔텔레키와 같은 신비한 개념을 가정할 필요가 없다는 것을 보여주었다. 그들은 유전자에는 구조유전자뿐 아니라 그것의 발현을 조절하는 제어유전자(operator)가 있다는 것을 발견했다. (제어유전자와 그것에 의해 제어되는 한 묶음의 구조유전자를 합쳐서 오페론이라고 부른다.) 이 제어유전자에는 통상 구조유전자의 발현을 억제하는 리프레서(repressor)가 붙어있다. 그러나 분해해야 할 물질이 들어오면 이것이 리프레서의 형태를 변형시켜 버리는데 이 때문에 리프레서가 오프레이터에서 떨어져 나가게 된다. 그러면 구조유전자의 전사(transcription)가 시작되고 그 물질을 분해하는 분해효소가 만들어진다. 카우프만은 이 아이디어를 형태형성의 유전자발현의 문제로 확장했다.
자콥과 모노는 이 작은 분자가 유전자를 켤 수 있다는 것을 발견했다. 리프레서 자체도 유전자의 산물이기 때문에 유전자들은 유전적 환류를 이루고 있어 서로를 켜고, 끌 수 있다는 것이 분명해졌다. 세포분화는 이러한 유전적 환류에 의해서 제어되고 있는지 모른다. 이러한 계를 잠깐 고찰해 보자. 유전자1이 유전자2를 억제하고, 유전자2가 유전자1을 억제한다면 이러한 계는 2가지 상이한 유전자 활성패턴을 갖게 된다. 첫 패턴에서는 1이 활성화되고 2가 억제된다. 둘째 패턴에서는 2가 활성화되고 1이 억제된다. 유전자 발현의 2가지 상이한 패턴으로 해서 이 유전적 환류가 2가지 상이한 세포유형을 만들어낼 수 있다. 각 세포는 동일한 유전적 회로의 選言的 패턴을 이루고 있다. 두 세포가 동일한 유전형을 가지면서도 상이한 조합을 통해서 상이한 형태를 발현시킨다… 그들의 작업은 세포분화가 어떻게 일어나는가를 시사하고 있다.
인간의 수정난 속의 10만 여 개의 유전자는 상호 연결된 네트워크를 형성하고 있으며 서로 상대를 켜고, 끔으로써 256가지의 상이한 세포형태를 발현시키는 것이 아닐까? 그런데 10만개로 만들어질 수 있는 가능한 연결의 수는 즉 개다. 이 가운데 250여 개만 발현한다는 것이 어떻게 가능한가? 카우프만은 우선 100개의 전구의 간단한 연결망을 만들어서 실제 이러한 일이 발생하는지를 실험해 보기로 했다.
……
그러나 이 “그 무엇”은 비물질적인 “생기”가 아니고 유전자들간의 상호작용이 만들어내는 자기촉매적 네트워크이다. 이 네 트워크는 주의 깊은 계획에 따라 만들어지는 것이 아니고 요소들간의 상호작용, 그리고 전체와 요소들간의 상호작용에 의해서 저절로 만들어지는 자생적 질서이다. 스스로 자기조직화 되어가는 이 네트워크는 범세계적으로 자기조직화 되어가고 있는 인터넷의 미래를 보여주는 것 같아 흥미롭다. (두렵다고 해야 할까?)
이제 우리는 더 원리적인 문제로 들어가 보자. 왜 상호연결의 네트워크가 만들어지면 상태의 수는 그것의 제곱근으로 축소되는 것일까? 다수 요소들간의 연결이 상호제약, 즉 대립적 제약(conflicting constraints)을 만들어내기 때문이다. 이 문제를 적합성 지형이라는 약간 다른 각도에서 살펴보자.
□ 적합성 지형과 NK모델
카우프만이 확인한 이 부존질서는 창조론과 진화론의 논점을 지양하면서 더 높은 차원에서 통합하고 있다. 그러한 의미에서 기본적 의도에서 베르그송의 “창조적 진화”와 그 맥을 같이하고 있다고 보여진다.
창조론자들의 진화론에 대한 비판의 핵심은 진화론자들이 주장하는 우연만으로는 복잡한 생명체가 만들어질 확률이 全無하다는 것이다. 이것은 부품들이 제멋대로 흩어져 있는 격납고에 허리케인이 불어 보잉 747기가 저절로 조립되었다고 주장하는 것처럼 우스꽝스럽다. 그러므로 생명이 가능하기 위해서는 어떤 식으로든 설계가 있어야 한다. 여기에 대해 진화론자들은 자연선택의 과정에서 질서가 나올 수 있기 때문에 특수설계를 전제할 필요가 없다고 주장한다. 즉 자기복제 과정에서 끊임없이 변이가 출현하고 있으며 그 가운데 환경과 적합한 것만이 살아남아 자신의 형질을 다음 세대에 전달할 수 있다. 환경에 적합한 것이 선택되고, 부적합한 것은 도태되는 과정이 바로 고도한 질서를 만들어내는 과정이라는 것이다. 말하자면 자연선택은 제멋대로 흩어져 있는 부품들에서 보잉747기를 조립할 수 있는 놀라운 기적을 행할 수 있다.
카우프만은 자연선택이 질서를 만들어내기 위해서는 어떤 선행조건이 필요하다고 보았다.그는 이것을 논증하기 위해서 1930년대 시웰 라이트(C.Wright)에 의해서 제안된 “적합성 지형”(fitness landscape)이라는 개념을 가져왔다. 여기서 적응은 언덕 오르기에 비유된다. 높은 언덕에 오르는 것은 적응도가 높은 것으로 간주된다. 경쟁이 시작되면 모든 종은 일제히 언덕 오르기를 시작할 것이다. 그런데 어떤 종의 언덕 오르기는 다른 종의 지형을 바꾸어 놓는다. 예컨대 개구리에 유전적 변이가 일어나 진뜩진뜩한 혀가 주어지면 그것은 파리를 쉽게 잡을 수 있어 개구리의 지형이 상승하고 이것이 파리를 언덕에서 끌어내려 파리의 적합성 지형을 바꾸어 놓는다. 이제 파리가 뱀에 어떤 질병을 옮기는 매개곤충이라고 하자. 파리의 지형도의 변화는 뱀의 적합성의 지형을 위로 끌어 올린다. 이 결과 뱀이 늘어 개구리의 적합성 지형을 도로 아래로 끌어내릴 것이다. 이처럼 한 종의 적합성의 정도는 다른 종의 적합성의 정도에 복잡한 방식으로 상호 의존하고 있다.
이것을 게놈내의 유전자들간의 상호작용에 적용시킨 것이 카우프만의 NK모델이다. 게놈내의 한 유전자의 상태는 다른 유전자의 상태에 복잡한 방식으로 상호 얽혀있기 때문에 앞서 개구리의 진뜩진뜩한 혀를 만든 유전자의 돌연변이는 게놈내의 다른 유전자에 영향-긍정적, 또는 부정적-을 줄지 모른다. 이처럼 유전자들간에 짝을 이루면서(coupling) 상호작용을 일으키는 것을 유전학자들은 “上位的 상호작용”(epistatic interaction)이라고 부른다. 이제 일반적으로 n개의 유전자가 k개의 다른 유전자로부터 영향을 받는 경우 만들어지는 적합성지형을 살펴보자.
카우프만은 k값을 변경시킴으로써 지형의 凹凸(ruggedness)과 봉우리의 수를 변경시킬 수 있다는 것을 발견했다. 즉 유전자들간의 상호작용을 증가시키면 대립적 제약들이 많아져서 많은 작은 국지적 봉우리를 가진 울퉁불퉁한 지형이 된다. 사실 상호간의 네트워크가 만들어지면 일방적 독주를 견제하는 대립적 제약이 불가피하게 나타난다. 앞서 개구리-파리-뱀의 네트워크를 보자. 자신에게 이익이 되는 것을 1로, 손해가 되는 것을 0으로 표시하자. 그러면 개구리가 끈적끈적한 혀를 개발했을 경우 1→0→1이 되고 이것은 다시 0→1→0,1→0→1…로 두 상태를 순환한다. 이 조그마한 네트워크에도 대립적 제약이 작용해서 어느 한쪽의 일방적 독주를 견제하고 있는 것을 볼 수 있다. (사실 유기체뿐만 아니라 복잡한 인공물도 설계상에 모순되는 디자인 기준에 직면한다. 전체설계 가운데 한 부분의 해결은 다른 부분에 대한 최적설계와 충돌한다. 그래서 상이한 하위문제들의 대립적 제약이 봉착하는 공통문제에 대한 타협 책을 찾아내어야 한다. 예컨대 튼튼한 자동차는 이상적이지만 속도나 연료의 소모량과 대립된다. 이 대립되는 디자인 기준은 설계상에 아주 풀기 어려운 최적화의 문제를 낳는다.) 이 모형에서 k는 1이지만 k값이 커지면 대립적 제약들이 늘어나서 k값이 적을 때 보이는 순환주기와 같은 질서는 볼 수 없고 복잡한 혼돈의 양상이 나타난다. 다시 말해서 수많은 작은 봉우리들로 된 울퉁불퉁한 지형으로 변한다.
지형구조와 적응적 진화간의 보다 선명한 상을 얻기 위해서 조종손잡이 k를 0으로 두고 시작해 보자. 아무런 상위입력도, 상호연결도 없기 때문에 대립적 제약도 없다. 이 지형은 하나의 봉우리가 우뚝 솟아있고 주위가 완만한 경사를 이루고 있는 일본의 후지산을 닮았다. 대립형질 1,0중 1이 더 적합한 것을 나타낸다고 하자. 그러면 (1111111111)은 전역적 최적값(global optimum)이다. 여기에는 국지적 봉우리들은 없다. 왜냐하면 다른 유전자 예컨대 (0001111111)는 변이를 통해서 0을 1로 뒤집음으로써 쉽게 전역적 최적값에 도달할 수 있기 때문에 국지적 봉우리로 고착되기 어렵기 때문이다. 더 나아가 각 인근의 게놈들 간에도 적합성에 있어서 큰 차이가 날 수 없는데 1에서 0으로 하나의 유전자를 변화시키는 것은 기껏해야 단지 1/n만큼 변화시키는 것에 지나지 않기 때문이다. 그래서 인근의 게놈들간의 적합성에는 근소한 차이들이 있을 뿐이다. 그래서 하나의 봉우리에 완만한 경사를 가진 지형이 만들어진다. 무작위로 된 유전형에서 시작한다고 하자. 그렇다면 유전자의 반은 1일 것이고, 반은 0일 것이다. 그러므로 봉우리까지의 예상되는 탐색거리는 n/2이다. 예컨대 (0101010101)의 경우 거리는 10/2 즉 5인데 5개의 0을 1로 변이시키면 최고봉에 도달할 수 있다. 한 단계 올라설 때 마다 거리는 하나씩 줄어든다. 최악의 유전형 (0000000000)에서 시작하더라도 언덕 오르기에 성공할 때 마다 한 단계씩 줄어들어 적응적 탐색은 어렵지 않게 최적값에 이를 수 있을 것이다.
그러나 이것은 N값이 비교적 작을 때 나타나는 효과이며 이 값이 커지면 개체군 전체의 변이율이 높아져서 덜 적합한 변이와 더 적합한 변이간의 선택적 차이가 개체군을 봉우리로 되돌아오게 하는 것 보다 더 빠른 속도로 봉우리 밖으로 흩어지게 만든다. 이것이 소위 “오류파국”(error catastrophe)인데 이 효과 때문에 이 지형상에서는 복잡한 고등유기체의 진화는 불가능하다.
보다 그럴듯한 모델을 만들기 위해서는 k값이 커져야 한다. k값을 그 최대값 N-1까지 돌려서 모든 유전자가 다른 유전자에 영향을 주는 경우를 살펴보자.k가 최대값이 되면 적합성 지형은 완전히 무작위 하게 된다. 어떤 하나의 유전자를 다른 대립형질로 바꾸는 것, 즉 0을 1로, 1을 0으로 바꾸는 것은 자신과 모든 다른 유전자에게 영향을 준다. 여기서는 북경에서의 나비의 날개짓이 뉴욕에 폭우를 몰고 오는 소위 “나비효과”(butterfly effect)를 일어난다. 여기서는 유리한 변이를 축적할 방법이 없다. 다음 단계에서 완전히 그 효과가 지워져 버리기 때문이다. 이 환경에서는 자연선택에 의한 질서의 구축은 불가능하다. 그래도 그것이 가능하다고 한다면 40억년의 기간 동안 매일 복권에 연속해서 당첨되는 극단적으로 좋은 운수였다고 말할 수 밖에 없다.
실제 지형은 후지산 지형처럼 단순하지도 않고, 또 그 반대로 완전히 무작위적인 것도 아니다. 모든 유기체는 k=0과 k=N-1사이의 어느 점에 위치한 지형 속에서 진화한다. 다윈의 진화론은 유전자의 무작위적 변이와 환경에 적합한 것의 선택에 기초하고 있다. 이 선택이 효력을 발휘하려면 k값이 0이 되어서도 안되고 그렇다고 k값이 너무 커서도 안 된다. 장기간의 컴퓨터시뮬레이션의 결과 카우프만은 k=2에 조율되어 있을 때 선택이 가장 효율적으로 작용한다는 것을 찾아내었다. 이것보다 낮으면 오류파국을 피하기 어렵고, 이것보다 높으면 대립적 제약이 커져 지형이 울퉁불퉁해져 국지적 봉우리에 갇히게 됨으로 전체의 적합성이 감소해가기 때문이다.
우리는 앞서 유전자 연결망에서 입력수가 2이면 끌개의 수가 상태수의 제곱근으로 축소되는 것을 보았다. 이것을 지금의 논의와 연관시켜 보면 이 적절한 수의 대립적 제약이 한편으로는 지나친 경직화를 피함으로써 오류파국을 막고 또 다른 한편으로는 지나친 유동성을 피함으로써 혼돈으로 빠지는 것을 막아주고 있다. 여기서 외부조건에 덜 민감한 끌개가 만들어지며 인간의 경우 의 가능한 경로 가운데 250여 개의 경로에 자신을 한정하고 있다. 지나친 경직성과 지나친 유동성의 경계선상에서 복잡계로서의 생명은 진화한다. 이것은 랭턴(C.Langton)이 제시한 “카오스의 가장자리”(the edge of chaos)의 또 다른 재발견임을 알 수 있다.
다윈주의자들은 자연선택 자체가 질서의 원천이라고 주장한다. 그러나 자연선택은 그것을 효율적으로 작동 가능하게 하는 어떤 지형이 주어져 있지 않을 경우 아무런 기능도 할 수 없다. 완만한 지형에서 선택의 기능은 변화를 위한 선택의 기능 보다는 변화의 저지를 위한 도태의 기능이 더 중요하다. 거기서 자연선택이 할 수 있는 것은 질서를 만들기 보다 혼돈을 저지하는 것이다. 요철이 극심한 지형에서는 선택은 축적되지 못하고 다음 단계에서 무효화되어 버리기 때문에 전혀 효력을 발휘할 수 없다. 자연선택에 진화가능성을 열어주는 이 특별한 지형은 그 자체 선택의 산물일 수는 없다. 자연 속에 내재해 있는 부존 질서 하에서만 자연선택은 그 효력을 발휘할 수 있다.
선택 그 자체만으로 선택이 잘 작동하는 그러한 종류의 지형에 적응한 유기체들을 만들 수 있고 유지할 수 있는지가 나로서는 아주 의심스럽다. 선택 그 자체만으로 진화가능성을 산출하고, 유지할 수 있다는 것은 전혀 분명하지 않다. 세포와 유기체가 선택이 작동할 수 있는 그러한 종류의 실재가 아니라면 어떻게 진화는 진화를 위한 발판을 얻을 수 있었을까?…
우리는 놀랄만한 가능성으로 되돌아오고 있다. 자기조직화가 진화적 능력의 전제조건이라는 것, 그것이 자연선택을 효율적 도구로 만들었다는 것이다. 그것이 점진적으로 진화할 수 있는 강인한 구조를 만들었다는 것이다. 왜냐하면 자생적 질서, 강인성(robustness), 용장성(redundancy), 점진성, 상호연관 된 지형 간에는 깊은 연관이 있기 때문이다. 용장성을 가진 계는 돌연변이에도 불구하고 아무런 영향을 받지 않거나 사소한 변화 만에 한정시키는 힘이 있다. 용장성은 점진적 변화를 산출하는 힘이다. 용장성의 또 다른 이름은 강인성이다. 그것은 많은 세세한 변화에 영향을 받지 않도록 하는 힘이다. 용장성의 또 다른 이름은 구조적 안정성이다. 접혀진 단백질구조, 조립된 바이러스의 구조, 부울리안 네트워크 등이 그것이다.
이 견해가 대략적으로 타당하다면 자기 조직화된 바로 그것이 전제됨으로 자연선택은 효율적으로 작동할 수 있다. 자기조직과 자연선택 간에는 어떠한 근본적 대립도 없다. 질서의 이 두 원천은 자연의 파트너이다. 게놈 네트워크는 혼돈의 가장자리에 가까운 질서의 영역에 있다. 그러한 네트워크는 쉽게 형성되는 질서, 부존질서의 일부이면서 다른 한편으로 구조적으로, 동력학적으로 안정되어 있다. 그래서 그것은 상호 연관된 지형상에 적응할 수 있으면서도 새로운 역할이 주어졌을 때 재구성될 수도 있다.
만일 자연선택이 자기조직적인 강인한 특성들을 이용해서 유기체를 만들었다면 우리는 뜯어 맞춘 임시방편적 분자기계가 아니다. 진화는 우연이 아니며 근저에 있는 질서의 표현이다. 우리는 예상된 존재이다. 그리고 우주는 우리의 집이다.
아래는 2014년 10월 6일 기사입니다~
(원문: 여기를 클릭하세요~)
생물과 무생물의 경계는 없다
네덜란드 이펜베르그의 모래 언덕에 바람이 부는 날이면 당신은 커다란 버스 크기의 조형물이 언덕을 천천히 올라가는 것을 볼 수 있습니다. 플라스틱 튜브와 나무, 돛이 복잡하게 얽힌 수많은 다리를 움직여 이동하는 이 물체는 마치 살아있는 듯한 느낌을 줍니다. 이들은 네덜란드의 예술가 테오 얀센의 작품입니다. 그의 웹사이트에는 이렇게 써 있습니다. “1990년부터 나는 새로운 종류의 생명체를 만들어야 겠다는 생각에 빠져있었습니다.” 그는 그의 작품들을 스트랜드비스트(strandbeest)라고 부릅니다. “나는 언젠가는 이들을 해변가에 무리지어 흩어놓고 싶어요. 그러면 이들은 자신만의 삶을 살게 되겠죠.”
대부분의 사람들은 스트랜드비스트가 어떤 감정을 불러일으키기는 하지만, 이들이 살아있는 것은 아니라고 말할 겁니다. 이들은 정교하고 아름답지만 생명을 가지지 않은 어떤 기계에 불과하다고 말이죠. 몇 달 전까지만 해도 나 역시 이런 생각에 동의했을 겁니다. 그러나 그 때는 내가 생명의 본질에 대한 진정한 통찰력을 가지기 전이었습니다. 이제 나는 스트랜드비스트가 곰팡이나 식물과같은 다른 생명체들과 전혀 다를 것이 없다고 생각합니다. 사실, 나는 어떤 것도 진정으로 살아있다고 생각하지 않습니다.
생명이란 무엇일까요? 이것은 과학이 답할 수 있는 문제가 아닙니다. 아리스토텔레스 이후 철학자와 과학자들은 생명의 보편적이고 정확한 정의를 찾기 위해 노력했습니다. 오늘날의 교과서에는 생명은 조직적이며, 성장하고, 번식하고, 진화하는 특성을 가지고 있다고 써있습니다. 그러나 이 정의에 맞지 않는 수많은 예외적 생명체들이 존재합니다.
예를 들어, 수정 결정은 매우 조직적이며, 성장하고, 자신의 형태를 충실히 복제합니다. 그러나 우리는 수정이 살아있다고 생각하지 않습니다. 또 어떤 컴퓨터 프로그램들은 번식하고, 짝을 짓고, 진화하지만 이들 역시 우리는 생명체로 여기지 않습니다. 반대로, 젤리 모양의 미생물인 완보동물(tardigrades)이나 아르테미아 새우는 수 년 간의 동면기간 동안 먹지도 않고, 성장하거나 형태를 바꾸지도 않음에도 불구하고, 우리는 이들을 살아있다고 생각합니다.
90년대 NASA의 과학자들은 다른 행성의 생명을 찾기 위해 생명을 임시로 정의해야 했습니다. 그들은 “진화능력을 갖추고 스스로 유지할 수 있는 시스템”을 생명이라고 정의했습니다. 물론 이 정의도 충분하지는 않습니다.
예를 들어, 자신의 DNA와 RNA를 단순한 단백질로 감싸고 있는 바이러스를 생각해 봅시다. 이들은 다른 세포에 침입해 자신을 복제할 수 있습니다. 또한 어떤 생명체보다도 빠르게 진화합니다. 그러나 생물학자들은 이들을 살아있는 생명체로 여길 것인지에 대해 아직 확실한 결론을 내리지 못하고 있습니다. NASA 에 참여했던 생물학자 제랄드 조이스는 바이러스는 “스스로 유지할 수” 없기 때문에 생명체가 아니라고 말합니다. 사실 바이러스는 자신이 감염시킨 다른 세포 안에서만 진화할 수 있습니다.
그러나 한편으로 생각해보면, 우리는 다른 기생 생명체들을 비록 이들이 홀로 살아갈 수 없다 하더라도 당연히 살아있는 것으로 여깁니다. 내장의 기생충이나 다른 식물의 수액을 빠는 덩굴식물, 거미를 감염시켜 죽인 후 그 사체에서 오렌지 색의 뿔을 키우는 곰팡이에 이르기까지 이들이 바이러스처럼 그 숙주에 번식과 진화를 의존한다는 것은 분명하지만, 이들은 살아있는 생물로 여겨집니다.
제랄드 조이스는 10년 동안 NASA와 함께 일한 이후, 자신들이 내렸던 생명의 정의를 더 위태롭게 만드는 한 실험과 마주쳤습니다. 그들은 실험실에서 때때로 서로를 복제하는 두 RNA 분자 쌍을 발견했습니다. 40억년 전, 지구가 원시 수프 상태일 때 이와 비슷한 RNA 들이 우연히 만들어졌을 것입니다. 이들은 바이러스보다 더 간단한 형태이지만 번식과 진화가 가능합니다. 조이스는 이들이 NASA 의 정의에 부합한다는 것을 인정했지만 이들이 살아있다고는 쉽게 말하지 못했습니다.
왜 이런 모순들이 발생하는 것일까요? 왜 과학자들은 생물과 무생물을 구분하는 것을, 그리고 바이러스가 생명체인지를 결정하기를 그렇게 어려워할까요? 그것은 그들이 실재하지 않는 어떤 개념을 정의하려 했기 때문입니다. 그것이 나의 결론입니다. 생명이란 그저 관념일 뿐입니다. 현실 세계에서 존재하고 구별되는 개념이 아닙니다.
내 말을 더 잘 이해하기 위해서는 우리가 가지는 마음속에 떠올리는 모형과 순수한 개념 사이의 차이를 구분하는 것이 필요합니다. 때때로 우리의 두뇌는 어떤 대상을 나타내는 표상을 만들어냅니다. 눈에 들어온 소나무에서 반사된 빛과 코를 자극하는 솔잎의 분자들의 자극들이 더해져 우리는 나무에 대한 기억을 만들게 됩니다. 뇌는 다른 대상에 대한 개념들 역시 이러한 관찰을 기반으로 만들어내며, 이는 세상을 인식하는 유용한 방법입니다. 우리가 “나무”라고 말할 때 우리는 순수한 개념으로의 나무를 생각합니다. 그러나 우리가 생각하는 “나무”는 우리의 마음속에만 존재합니다. 세상에 우리가 나무라고 뭉뚱거려 표현하는 수십억의 식물들이 존재하는 것은 사실입니다. 당신은 식물학자들이 나무에 대한 명확한 정의를 가지고 있을 것이라 생각할지 모르지만, 이는 사실이 아닙니다. 어떤 식물이 나무(tree)인지, 관목(shrub)인지를 구별하는 것은 때로 매우 어렵습니다. 왜냐하면, “나무”와 “관목”은 식물이 가진 고유의 특성이 아니라 우리가 그들에게 붙인 이름에 불과하기 때문입니다.
“생명(life)”도 이와 비슷합니다. 우리가 어떤 대상들을 살아있는 것과 살아있지 않은 것으로 구분하는 것은 종종 유용할 수 있지만, 그러한 구분은 사실 우리의 마음 속에만 존재하는 것입니다.
사실 생명을 정의하는 것은 쓸데없는 일일 뿐 아니라 그 대상이 어떻게 작동하는지를 이해하는 데 불필요하기까지 합니다. 가장 기초적인 수준에서 유일하게 의미있는 것은 그 대상을 이루는 원자들과 다른 입자들이 어떻게 구성되어 있는가 하는 것입니다. 복잡도의 척도에는 물 분자 하나처럼 극히 간단한 대상에서부터 개미집처럼 놀랄만큼 복잡한 대상들이 존재합니다. 생명의 특징이라 여겼던 대사, 번식, 진화와 같은 활동들은 이 복잡도 척도상의 여러 다른 수준에서 나타나는 현상들입니다. 생명의 경계로 딱 떨어지는 경계선은 존재하지 않습니다.
우리가 살아있지 않다고 생각하는 어떤 것들이 때로 우리가 생명체만의 특성으로 여기는 그런 활동들을 할 수 있습니다. 반대로 우리가 살아있다고 여기는 것들이 그런 특성을 갖추지 못하기도 합니다. 우리는 지금까지 이들을 단 하나의 기준, 곧 살아있는 것과 살아있지 않은 것으로 구분하려 했으며, 현실에서 아무런 의미없는 경계선을 찾으려 노력했습니다.
그런 것은 존재하지 않습니다. 우리는 생명이란 개념이 그저 우리의 편의를 위해 만들어진 실용적인 개념일 뿐이며, 우주의 실재를 반영하는 것이 아니라는 사실을 인정해야 합니다.
이를 인정하는 것은 여러모로 우리의 생각을 자유롭게 만들어 줍니다. 우리는 더 이상 얀센의 작품들이 살아있는 것인지를 고민할 필요가 없습니다. 스트랜드비스트가 우리를 매혹시키는 이유는, 이들이 “살아있는 것들”이 우리를 매혹시키는 그 특성들을 정확히 가지고 있기 때문이며, 이것은 이들이 매우 복잡하기 때문이고, 바로 그 복잡함 속에 아름다움이 존재하는 것입니다.
스트랜드비스트의 돛이 바람에 의해 흔들릴 때, 다리들은 규칙을 가지고 굽혀지고 펴지며, 이 물체의 움직임은 시작됩니다. 나는 스트랜드비스트의 의지와 집요함을 느낄 수 있습니다. 이 장엄한 존재가 살아있는가 그렇지 않은가는 중요하지 않습니다. 그저 지켜볼 뿐입니다.
(뉴욕타임즈)
아래는 2019년 9월 2일 기사입니다~
(원문: 여기를 클릭하세요~)
생명이란 무엇인가
삶과 죽음, 그 경계에 대하여
![[shutterstock]](https://dimg.donga.com/ugc/CDB/WEEKLY/Article/5d/68/b6/22/5d68b6221271d2738de6.jpg)
[shutterstock]
삶과 죽음, 그 경계선은 얼마나 뚜렷합니까. 갈색 곱슬머리의 아일랜드 과학자가 숨죽인 강연장에서 첫 마디를 열었다. 삶과 죽음에 대한 것이라니. 노년의 철학자가 꺼낼 법한 이야기가 푸른 눈빛의 젊은 과학자에게서 나오자 청중은 술렁이기 시작했다. 그는 편안하게 말을 이었다.
캐나다 북부로 가면 삶과 죽음의 경계에 있는 송장개구리(학명 Rana sylvatica)를 만날 수 있다. 캐나다 송장개구리는 아주 특별한 재주가 있다. 모두 알고 있듯이, 개구리는 변온동물이어서 주위 온도에 따라 자신의 체온 역시 변한다. 문제는 캐나다 송장개구리는 적당히 낮은 온도가 아니라 매우 추운 환경에서 산다는 것이다. 기온이 영하로 떨어지면 송장개구리는 얼기 시작한다. 제일 먼저 다리가 얼고, 그다음은 머리와 가슴이 언다. 그리고 결국 심장까지 얼어버린다.
얼지 마, 죽지 마, 부활할 거야
![캐나다 송장개구리. [shutterstock]](https://dimg.donga.com/ugc/CDB/WEEKLY/Article/5d/68/b6/5d/5d68b65d11e5d2738de6.jpg)
캐나다 송장개구리. [shutterstock]
여기서 언다는 것은 비유가 아니다. 실제로 심장박동이 없고, 맥박도 없다. 몇 주간 캐나다 송장개구리는 우리 기준으로 죽은 상태로 지내는 것이다. 하지만 봄이 되면 놀라운 일이 벌어진다. 개구리의 심장은 천천히 다시 움직이고, 날씬한 다리로 뛰어다니기 시작한다. 어떻게 이런 일이 가능할까. 마치 죽음에서 부활하는 것과 비슷한데 말이다.
인간의 경우 몸속 세포가 얼기 시작하면 수분이 세포에서 빠져나가 탈수 상태가 된다. 세포 주변이 얼고 더는 수분이 남아 있지 않게 되면 세포는 부서지고 죽게 되는 것이다. 반면 송장개구리는 세포가 얼기 시작하면 간에 저장돼 있던 녹말이 포도당으로 분해되고, 이 포도당이 혈관을 타고 심장을 포함해 주요 장기 및 세포 속 수분과 결합해 마치 설탕물처럼 만들어준다. 설탕물은 물보다 어는점이 낮다. 그래서 세포가 완전히 얼지 않는 것이다. 천연 내한제를 몸속에 보유하고 있는 셈이다. 추위를 피하지 않고 받아들인 캐나다 송장개구리는 삶과 죽음의 경계를 허물어뜨린다. 얼어 죽는 대신 얼어 사는 것이다.
모든 생명체의 근원, 세포
![퍼거스 매콜리프 교수. [TEDx Talks 유튜브]](https://dimg.donga.com/ugc/CDB/WEEKLY/Article/5d/68/b6/7c/5d68b67c1b4ed2738de6.jpg)
퍼거스 매콜리프 교수. [TEDx Talks 유튜브]
훌륭한 발표를 마친 그의 이름은 퍼거스 매콜리프(Fergus McAuliffe). 아일랜드의 유니버시티 칼리지 코크의 환경과학자인 그는 사람의 생명을 살리기 위한 냉동장기 연구를 하고 있다. 삶과 죽음의 경계에서 죽음으로 가는 시간을 최대한 늦추기 위한 방법을 찾고 있는 것이다. 이뿐 아니라 수많은 과학자도 비슷한 연구를 하고 있다. 그만큼 생명이 소중하기 때문이다. 그렇다면 여기서 중요한 생명이라는 것, 그 정체는 무엇일까.
생명이 있는 것을 우리는 생물이라 부르며, 그렇지 않은 것은 무생물로 분류한다. 무생물은 정리되지 않은 방 안과도 같다. 언제부터 쌓였는지 모를 잡동사니와 각종 쓰레기가 무분별하게 어질러져 있다. 생물은 여기서 약간의 규칙과 노력을 통해 어느 정도 정리된 것이라고 볼 수 있다.
지구상 모든 살아 있는 생물은 ‘세포’라는 아주 작은 단백질 기반의 부품으로 이뤄져 있다. 세포는 테두리에 벽을 갖고 있어 주변 환경으로부터 자신을 보호하며, 스스로 반응하고 성장하면서 진화의 주체가 되기도 한다. 그렇다면 세포야말로 생명의 근원이자 시작이 아닐까.
재미있는 사실은 세포가 생명의 가장 보편적 증거라 해도, 세포를 구성하는 모든 것은 전혀 살아 있지 않다는 점이다. 세포는 살아 있다. 허나 세포를 구성하는 각 부분에서 수백만 종류의 화학반응이 오케스트라처럼 복잡하게 일어나긴 해도 정작 세포의 어느 부분도 살아 있지는 않다. 좀 더 보편적으로 들어가면 생물은 죽어 있는 모든 것으로 이뤄진 세포의 동력기관이라고 볼 수도 있다. 무생물로 이뤄진 부품이 결합해 하늘을 나는 비행기가 되거나 꽤 빠른 자동차가 되는 것처럼 말이다.
결론적으로 살아 있는 모든 것은 죽는다. 다행히 여기서 죽음이란 우리가 생각하는 것처럼 절망적이지 않다. 이미 생명체를 구성하는 모든 것은 확실한 죽음에 이른 상태며, 그저 원래 상태로 잘게 분리되는 것이기 때문이다. 하지만 단순히 분리되는 것만으로 끝나지는 않는다. 살아 있는 생명체의 최종 목표는 다시 새로운 개체를 만들어내 영원한 죽음을 막는 것이기 때문이다.
생명체 가운데 운 좋게 가장 우수한 설계도에 따라 만들어진 생명체만 살아남게 되고, 그 설계도는 다양하게 변형돼 다음 개체에 다시 옮겨진다. 우리는 이 유전자 정보가 포함된 설계도를 DNA라 부르며, 살아 있다고 착각하는 순간은 단순히 정보를 옮기는 과정에 불과하다.
그렇다면 계속 옮겨지는 DNA야말로 생명이라고 볼 수 있을까. 설계도는 생명체에게 반드시 필요하며, 생존에 매우 중요한 요소다. 하지만 DNA 스스로는 아무것도 할 수 없다. 과연 생명이라는 것이 어디서 시작하는지, 어느 시점부터를 생명체로 볼 수 있는지는 확실하지 않다. 한 가지 확실한 점은 살아 있는 존재만 뭔가 특별히 신비로운 재료로 구성된 것이 아니라, 살아 있지 않은 것을 이루고 있는 똑같은 물질로 만들어졌다는 사실이다.
살아 있거나 혹은 죽어 있다는 착각
![애벌레가 나비로 환골탈태할 때 남겨진 고치는 무생물이지만 변태과정에서 생명의 일부이기도 했다. [shutterstock]](https://dimg.donga.com/ugc/CDB/WEEKLY/Article/5d/68/b6/e8/5d68b6e813cdd2738de6.jpg)
애벌레가 나비로 환골탈태할 때 남겨진 고치는 무생물이지만 변태과정에서 생명의 일부이기도 했다. [shutterstock]
생명의 근원이나 증거를 찾는 것은 너무 어려우니 질문을 조금 바꿔보자. 만약 이 우주의 모든 것이 전부 같은 물질로 구성돼 있다면 생명체라는 것이 존재할까. 과연 생명이라는 것을 별도로 구분할 필요가 있을까. 모두 죽어 있는 것일까, 아니면 반대로 전부 살아 있는 것일까. 우리를 구성하는 모든 것이 한 번도 살아 있은 적이 없었기에 결코 죽을 수 없는 불로불사(不老不死)라고 볼 수도 있을까.
생명이란 무엇인가. 그럼 정의란 무엇이며, 행복이란 무엇인가. 마치 무엇인지 알려주려고 한 것처럼 제목부터 직설적으로 묻는 책들에는 공통점이 있다. 결론은 무엇인지 모른다는 것이다. 미국 정치철학자 마이클 샌델 교수의 책은 셀 수 없을 만큼 많이 팔렸지만, 결코 정의라는 것은 책 한 권으로 넘어갈 수 있는 개념이 아니다. 당연한 이야기지만, 이스라엘 최고 랍비로 불리는 하임 샤피라 역시 행복에 대해 수학공식처럼 알려주는 책을 쓴 것이 아니다. 우리는 무엇인가를 알려주는 책을 통해 이게 무엇인지 쉽게 정의할 수 없다는 것을 경험적으로 터득할 수 있을 뿐이다.
생명도 마찬가지다. 이 짧은 글로 생명이 무엇인지를 알아내는 것은 불가능에 가깝다. 삶과 죽음에 대한 명확한 정의도 내릴 수 없다. 그저 우리는 생명이며, 삶과 죽음의 경계에서 계속 살아가고 있을 뿐이다. 우리는 계속 살아야 한다. 생명이란 무엇인지 끊임없이 생각하며.
아래는 2023년 7월 18일 뉴스입니다~
(원문: 여기를 클릭하세요~)
‘분자 배터리’가 생명의 에너지를 흐르게 한다
(9) 생명의 대사 작용 원리

에너지를 이용할 줄 아는 능력이야말로 생명의 핵심이다. 픽사베이
이번 칼럼부터는 예고대로 지구 생태계에 대한 이야기로 돌아 가보자. 기억을 되살리기 위해 이전 내용을 요약하면 다음과 같다.
(1) 무질서의 정도 즉 엔트로피의 증가는 자연 법칙이다, (2) 엔트로피 증가는 죽음을 의미한다, (3) 죽음을 거스르기 위해서는 에너지가 필요하다, (4) 생명을 구성하는 고분자에는 연결 에너지가 존재한다, (5) 생태계의 생물들은 생존 경쟁을 통해 에너지와 자원을 순환시킨다. 이번 칼럼에서는 생명 에너지에 대해 구체적으로 알아볼 것이다.
우리가 계속 이야기하고 있는 생명은 무엇일까? 이는 의외로 정답을 찾기 어려운 질문이다. 종교, 학문, 시대적 배경 그리고 개인적 관점에 따라 생명은 다양하게 정의된다. 하지만 현대 생물학 지식을 바탕으로 다양한 정의들을 정리하면 대사(metabolism)와 복제(replication)가 생명의 핵심으로 요약된다. 에너지를 이용하는 대사 능력, 그리고 자손을 복제하는 능력, 이 둘 다 가능해야 “생명을 가진” 생물로 분류된다. 이 가운데 복제에 대한 이야기는 다음으로 미루고 지금은 대사를 먼저 알아보자. 대사는 생명이 에너지를 이용하는 포괄적 과정으로, 두 개의 상반된 과정이 포함된다. 하나는 생체 고분자를 분해하는 이화(catabolism), 또 하나는 그 반대로 생체 고분자를 합성하는 동화(anabolism) 과정이다.
이화 과정에서는 고분자가 분해되면서 연결이 품고 있던 에너지가 방출된다. 반대로 동화 과정에서는 이화과정에서 얻은 저분자를 결합시켜 고분자를 합성해낸다. 고분자가 합성된다는 것은 원자들이 규칙적으로 연결된다는 것이다. 질서가 부여된다는 것은 엔트로피 감소를 의미한다. 즉 동화가 바로 죽음을 거스르는 과정이며 에너지가 소모되는 과정이기도 하다. 그럼 이 에너지는 어디에서 왔을까? 이화 과정에서 방출된 에너지를 이용한다.
그런데 이화와 동화는 같은 위치에서 일어나지 않는다. 따라서 이화에서 방출된 에너지를 동화에 효율적으로 전달해주는 수단이 필요하다. 만약 배터리가 없다면 핸드폰 사용이 얼마나 불편할지 상상해보라. 항상 전원 콘센트에 연결해야 한다면 핸드폰은 쓸모없는 물건이 된다. 무선 통신 기술과 함께 어디서나 에너지를 공급하는 배터리 덕분에 편리한 핸드폰이 탄생한 것이다. 대사에서도 배터리 역할을 하는 아데노신 삼-인산(Adenosine Tri-Phosphate, ATP)라는 분자가 존재한다. 이 생체 배터리가 없었다면 이화와 동화는 같은 위치에서만 일어나야 했을 것이고, 복잡한 생물은 존재할 수도 없었을 것이다. 생소한 화학 용어가 거슬리더라도 ATP는 그만큼 중요하니 꼭 기억해두자,
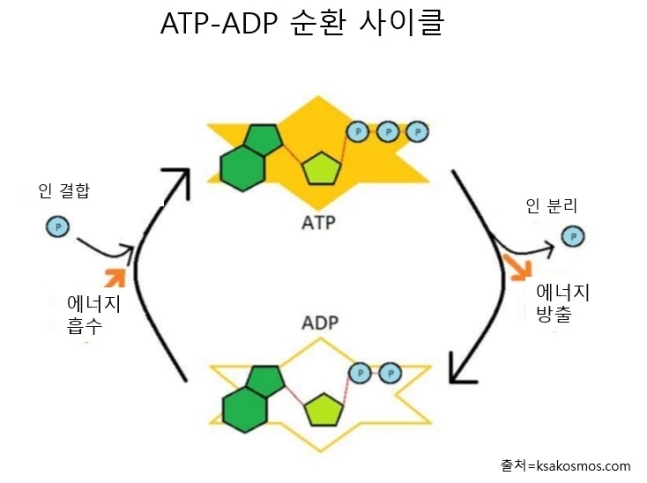
ADP와 ATP는 이화와 동화를 오가며 에너지를 전달하는 중요한 역할을 수행한다. 픽사베이
ATP와 ADP의 에너지 순환 시스템
ATP의 구조나 작동 원리를 자세히 알 필요는 없고, 원소 인을 뜻하는 P가 중요한 역할을 한다는 것만 알아두자. 이 인의 결합에는 많은 에너지가 저장된다. 인이 결합되면 충전, 떨어지면 방전이다. 충전된 상태는 ATP, 떨어진 상태는 ADP가 된다(ATP는 인이 세 개, ADP는 두 개 붙어있다는 뜻이다). 대사의 이화 과정에서 방출된 에너지는 방전된 ADP를 ATP로 충전하는 데 이용된다. 그리고 동화 과정에서는 충전된 ATP를 방전시켜 고분자 연결에 필요한 에너지를 얻게 된다. 생성된 고분자는 더 낮은 엔트로피와 더 많은 결합 에너지를 갖게 된다. 그리고 방전된 ADP는 다시 이화 과정으로 가서 충전된다. 이렇게 ADP와 ATP는 이화와 동화를 오가며 에너지를 전달하는 중요한 역할을 수행한다.
그런데 이런 복잡한 과정을 거쳐 고분자를 분해해서 다시 합성하는 것이 비효율적이라는 생각이 들 수도 있다. 하지만 분해되는 고분자는 먹이이고 합성하는 고분자는 자기 몸의 구성 성분이라는 차이가 있다. 예를 들어 새끼 고양이를 집안에서만 기른다고 생각해보자. 사료와 물만 먹이면 쑥쑥 자라서 새끼를 놓는다. 사료 속 고분자들이 분해되어 고양이의 형태로 재합성이 되고 최종적으로 자기 복제까지 일어나는 것이다. 이 모든 과정이 대사를 통해 진행된다.

웨이트 트레이닝은 단백질 가루를 근육으로 변환시키는 과정이다. 픽사베이
근육 운동으로 본 에너지의 ‘이화와 동화’
좀 더 실질적인 예를 들어보자. 바디빌딩의 목적은 근육을 만드는 것이다. 단백질 파우더를 섭취하면 이화를 통해 기본 구성 성분인 아미노산으로 분해된다. 그리고 동시에 포장에 표시된 칼로리의 에너지가 얻어진다. 물론 실제 인체에서는 소화 흡수 대사라는 여러 과정이 복잡하게 연결되지만 여기서는 단순화해서 생각하자. 그 다음 근육 세포의 한계까지 열심히 웨이트 트레이닝을 하면 해당 근육에서 근섬유 단백질이 더 필요하다는 자극을 받는다. 그럼 근원섬유를 구성하는 마이오신과 액틴 단백질의 동화가 증가하여 근육의 부피가 커지게 된다. 입력과 출력만 따져보면 단백질 가루가 근육으로 변환된 것이다.
인체에서 동화와 이화의 균형은 호르몬에 의해 정교하게 조절이 된다. 만약 근육의 생성에 충분한 칼로리가 음식을 통해 공급되지 않으면 인체의 에너지 저장고인 지방세포가 이화되어 에너지가 공급된다. 살이 빠지는 것이다. 만약 지방도 충분하지 않으면 오히려 근육이 이화되기 시작한다. 바디 빌더가 가장 무서워하는 단어인 ‘근손실’이 일어난다. 반대로 섭취된 칼로리만큼 운동을 하지 않으면 남는 에너지는 지방으로 변환되어 저장된다. 살이 찌는 것이다. 따라서 섭취한 칼로리와 운동량의 균형을 잘 맞춰야 원하는 부위에 근육이 제대로 만들어진다. 이것이 어렵다고 인위적으로 호르몬을 투여해 근육 동화를 비정상적으로 촉진하는 경우가 있다. 이런 약물이 바로 운동선수에게 금지 약물인 아나볼릭(동화) 스테로이드다. 이런 행위는 단기적으로는 큰 효과를 얻을 수 있지만, 장기적으로는 정교하게 조절되는 인체의 대사 균형을 교란시켜 심각한 부작용을 일으킨다.
고양이는 사료를, 사람은 음식을 먹어 동화에 필요한 에너지와 재료를 얻는다. 이를 자연 생태계에서 보면 사료와 음식은 피식자, 고양이와 사람은 포식자이다. 먹고 먹히는 관계가 차례로 연결되면 생태계의 먹이 사슬 혹은 피라미드가 만들어진다. 그런데 동화와 이화가 아무리 정교하게 일어나도 대사의 에너지 효율이 100%가 될 수 없다. 거기에 도망가고 잡기 위해 소모되는 에너지를 생각하면 계속 순환되는 생태계 에너지는 극히 일부에 불과하다. 따라서 에너지가 지속적으로 추가 투여되지 않으면 생태계는 죽어가게 된다. 즉 생태계에는 최초의 동화를 계속 수행해 에너지를 공급하는 구성원이 필요하다.

식물은 엽록체, 동물은 미토콘드리아가 에너지 공장이다. 픽사베이
식물과 동물의 에너지 공장, 어떻게 다른가
생태계를 가득 채우고 대사의 사슬을 구성하는 생물은 식물과 동물로 크게 분류할 수 있다. 식물과 동물의 차이는 무엇일까? 동물은 움직이는 것, 식물은 움직이지 않는 것이라는 생각이 먼저 떠오른다. 하지만 이보다 더 근본적 차이는 세포 내에서 ATP 충전을 담당하는 에너지 공장의 종류가 다르다는 것에 있다. 식물은 엽록체, 동물은 미토콘드리아가 에너지 공장이다. 미토콘드리아에서는 이화를 통해서만 충전이 일어나지만, 엽록체에서는 빛으로 ATP를 바로 충전할 수 있다.
따라서 생태계에서 최초의 동화작용은 엽록체를 가진 식물이 시작한다. 식물은 빛과 물과 이산화탄소만 있으면 탄수화물 동화를 지속하며 자란다. 단순한 원자들을 탄수화물이라는 고분자로 전환시키기 위해서는 연결 에너지가 필요하다. 식물은 이 에너지를 빛에서 얻는 특별한 능력을 가지고 있는데 이것이 광합성이다. 엽록체가 없는 동물은 다른 생물을 잡아먹어야 이화작용을 통해 에너지를 얻을 수 있다. 이때 에너지 이용 효율을 높이는 것이 미토콘드리아다. 이화 없이 에너지 충전이 가능한 식물은 독립 영양 생물, 이화를 통해서만 에너지 충전이 가능한 동물은 종속 영양 생물로 분류한다.

1억5천만km 떨어진 태양에서 온 빛 에너지가 지구 생명 에너지의 근원이다. 픽사베이
모든 것의 시작점은 태양 에너지
광합성이 지구 생태계에 에너지가 투입되는 유일한 수단이다. 그럼 광합성에 필요한 빛은 어디에서 왔을까? 이글거리는 태양은 1억5천만km 떨어진 지구에 빛 에너지를 전달한다. 태양 깊숙한 중심부에서는 두 개의 수소 원자핵이 하나의 헬륨 원자핵으로 융합되고 있다. 그런데 수소 원자핵 두 개의 질량보다 헬륨 원자핵 한 개의 질량이 더 작다. 에너지는 질량에 빛의 속도를 두 번 곱한 것과 같다는 아이슈타인의 유명한 공식에 따라, 사라진 질량은 에너지로 전환된다. 핵융합 과정에서 사라지는 질량은 아주 미미하지만 빛의 속도라는 엄청나게 큰 상수 값을 한 번도 아닌 두 번이나 곱하기 때문에 어마어마한 에너지로 전환된다. 태양 중심부에서 수소 폭탄이 계속 터지는 상황이라 생각하면 된다.
태양 중심부의 핵융합이 만들어내는 막대한 에너지는 빛으로 방출된다. 관찰 방법에 따라 빛은 파동이 되기도 하고 입자가 되기도 하는 이중성을 가지고 있다. 파동 형태이면 전자기파로, 입자 형태이면 광자로 불린다. 그리고 이 전자기파의 특정 파장은 우리 눈으로 관찰이 가능한 가시광선이다. 태양의 심부에서 방출된 빛이 태양 표면까지 도달하는 데는 십만년이 걸린다. 하지만 표면을 떠난 빛은 우주를 가로질러 8분 뒤면 지구에 도달한다.
지구에 도달한 대부분의 빛 에너지는 복사열로 흡수된다, 복사열은 바닷물과 육지를 데워서 지구 온도를 생명활동에 적절한 수준으로 유지해준다. 구역에 따라 흡수된 복사열의 차이는 비, 바람, 태풍 같은 기상 현상을 일으킨다. 하지만 생명은 태양이 보내주는 빛 에너지를 바로 이용할 수 없다. 생명 에너지로 변환되어야 한다. 마치 태양광 발전 패널을 통해 빛이 전기 에너지로 변환되는 것처럼, 식물은 빛을 생명 에너지로 전환시킨다. 여기서 생명 에너지는 추상적 개념이 아니다. 생명을 구성하는 특정 고분자들의 연결에 존재하는 화학 에너지가 생명 에너지이다.

대부분의 식물은 붉은색 파장의 빛(광자)을 흡수하고 나머지는 반사한다. 가시광선에서 붉은색만 흡수되고 반사되면 초록색이 된다. 픽사베이
대부분의 식물이 초록빛을 띠는 이유
지구는 자전을 통해 낮과 밤의 질서를 만든다. 이를 통해 표면의 식물들이 골고루 빛을 쪼이도록 해준다. 대부분의 식물은 붉은색 파장의 빛(광자)을 흡수한다. 그리고 나머지 색은 반사한다. 가시광선에서 붉은색만 흡수되고 반사되면 초록색이 된다. 즉 초록색은 광합성의 증거인 셈이다. 태양이 뜨거워질수록 식물의 초록은 더 깊어진다. 시인들이 봄의 신록을 노래하고 한여름의 깊은 초록에서 강렬한 생명이 느껴지는 것은 지극히 생물학적 반응이기도 하다. 계절이 바뀌면 광합성은 더 이상 동작하지 않는다. 붉은색을 흡수하지 못하는 잎은 초록을 잃어버리고 떨어지게 된다.
식물은 빛 에너지를 이용해 생태계의 가솔린이라 할 수 있는 탄수화물을 합성한다. 이 광합성은 빛을 이용하는 명반응과 이용하지 않는 암반응 두 단계를 거친다. 명반응은 태양전지에서 발생된 전기 에너지를 배터리에 충전시키는 것을 상상하면 된다. 이 과정에서는 빛 에너지를 흡수해 생태계의 배터리인 ATP를 충전시킨다. 암반응에서는 이산화탄소와 물을 재료로 탄수화물을 합성하게 된다. 이산화탄소의 탄소 원자를 분리해 생체 고분자로 재결합시키는 것이다. 이산화탄소와 물이라는 분자보다 탄수화물이라는 고분자가 가진 연결 에너지가 훨씬 크기 때문에 암반응에서는 에너지가 투입되어야 한다. 이때 명반응에서 충전된 ATP를 이용한다. 만들어진 탄수화물은 식물의 관을 따라 순환하며 줄기가, 뿌리가, 열매가 된다. 풀이나 나무가 자라는 속도를 보면 광합성이 얼마나 활발하게 일어나고 있는지 짐작이 가능하다.
식물이 독립 영양 생물이라면, 스스로 에너지를 생산하는 능력이 없는 모든 동물은 종속 영양 생물이다. 생태계에서 식물은 생산자, 동물은 소비자다. 그리고 동물은 다시 초식, 육식, 혹은 잡식으로 분류된다. 초식 동물의 경우 깨어있는 동안 거의 모든 시간을 풀을 먹고 소화하는 것에 투자해야 한다. 필요한 에너지와 생체자원을 얻기 위해서는 그만큼 많은 양의 식물을 섭취해야 하기 때문이다. 반면 육식성 동물은 에너지 효율이 초식동물보다 뛰어나다. 다른 동물을 잡아먹으면 더 많은 에너지를 효율적으로 획득할 수 있기 때문이다. 하지만 가만히 움직이지 않는 식물을 먹는 것과 죽기 살기로 도망치는 먹이를 잡아서 먹는 것은 난이도 측면에서 비교가 되지 않는다. 육식동물에겐 지능이 생존 능력이라는 의미다. 그래서 초식 동물보다 육식 동물의 평균적인 지능이 더 높다.
이런 에너지 흐름에만 식물과 동물의 연결고리가 있는 것은 아니다. 물, 이산화탄소, 빛이 필요한 광합성 과정이 끝나면 산소가 부산물로 배출된다. 동물은 호흡을 통해 산소를 받아들여 세포의 미토콘드리아에서 많은 ATP가 충전되도록 한다. 그리고 부산물로 이산화탄소를 배출한다. 동물이 배출한 이산화탄소는 식물의 기공을 통해 흡수되어 광합성의 재료로 쓰인다. 이런 이산화탄소와 산소의 순환 고리도 생태계에 존재하는 동식물 공생의 큰 축이다.

모든 생명은 식물에 의존하고 있다. 따라서 기후 변화 최악의 시나리오는 식량 위기다. 픽사베이
식물은 지구 생태계의 주춧돌
지구 생태계의 수많은 생명은 직접적이든 간접적이든 결국 태양 빛에 의존해서 살아가는 셈이다. 만약 화산 폭발이나 운석 충돌로 지구 대기가 먼지로 가득차서 식물에 빛이 차단된다면 생명 에너지 생산이 중단된다. 생태계에 추가적인 에너지의 공급이 없으면 생명을 유지하는 결합에너지도 부족해진다. 엔트로피는 증가하게 되고 생태계는 서서히 죽어가게 된다. 대멸종이 일어나는 것이다. 식물이 살지 못하면 동물도 살 수 없다.
요즘 기후 문제가 점점 커지고 있다. 기후 변화로 해수면이 상승하고 폭염, 홍수, 가뭄 같은 기상 이변이 점점 자주 발생한다. 하지만 더 무서운 것은 식물의 서식 환경 변화다. 우리나라도 점차 아열대 기후로 변하고 있다. 동물은 기온이 변하면 더 시원하거나 따듯한 곳으로 움직이면 된다. 하지만 움직일 수 없는 식물은 자라는 곳의 기후(온도와 강우량)가 변하면 죽어버린다. 현재 인류의 주식인 곡물은 소위 곡창 지대로 불리는 특정 지역에서 효율적으로 자라는 품종으로 개량되어 획일화되어 있다. 따라서 곡창 지대의 기후가 조금만 변해도 식량 생산은 타격을 받는다. 인류 문명이 아무리 발달해도 사람은 종속 영양 생물이다. 따라서 기후 변화 최악의 시나리오는 식량 위기다. 특히 우리나라처럼 식량자급률이 낮은 경우는 더 치명적인 시나리오이다. 밥 한 공기에 10만원, 라면 한 봉지에 20만원인 세상은 얼마나 끔찍할까. 고기를 대신 먹으면 된다고 생각하지 말기 바란다. 가축이 먹는 사료들도 대량 생산된 곡물이다. 다시 강조하지만 생태계의 모든 생명은 식물에 의존하고 있다.
이번 칼럼에서는 태양과 식물에 의존하는 생명 에너지를 대사를 중심으로 이야기하였다. 그런데 뭔가 이상하지 않은가? 어떻게 지구 생태계의 다양한 생명들이 품고 있는 고분자와 에너지가 서로 호환되고 순환이 되는 것일까? 그 이유는 현재 생태계의 모든 생물이 하나의 세포에서 기원했기 때문이다. 다음 칼럼에서는 LUCA(Last Universal Common Ancestor)라 불리는 생명의 공통 조상 세포에 대해 알아보기로 하자.
아래는 2023년 7월 22일 뉴스입니다~
(원문: 여기를 클릭하세요~)
세계 최초 ‘세포 지도’ 완성…장기와 세포 상호작용 확인

국제학술지 ‘네이처’는 이번주 표지로 3D 컴퓨터 모델링으로 구현한 장기들의 모습을 실었다. 팥 두개를 붙여놓은 듯한 신장을 비롯해 다양한 장기가 표현됐다. 표지에는 ‘몸의 그림(BODY IMAGE)’이란 문구가 적혀 있다.
마이클 스나이더 미국 스탠퍼드대 의대 교수 등이 참여한 국제공동연구팀인 ‘인간생체분자 아틀라스 프로그램(HuBMAP) 이니셔티브’는 인간의 장, 신장, 태반에 존재하는 세포들의 분포와 역할을 입체적으로 확인한 연구 결과를 19일(현지시간) 네이처에 발표했다. 단일 세포 수준에서 세포들의 정확한 분포와 역할을 확인한 세계 최초의 ‘세포 지도’다. 2018년 시작된 HuBMAP은 인체의 모든 세포가 어떻게 배열되는지 지도로 나타내는 것을 목표로 한다.
세포 지도는 질병 치료의 중요한 단서가 된다. 인체 곳곳에 존재하는 세포는 각종 장기나 조직과 상호작용하면서 신체 구성요소가 어떤 기능을 할지 결정하기 때문이다. 하지만 각 세포가 우리 몸속에서 어떻게 분포하고 작용하는지는 구체적으로 알려지지 않았다.
연구팀은 소화에서 면역 체계 형성에 이르기까지 다양한 기능을 하는 장기인 장에 주목했다. 20명의 기증자로부터 소장과 대장을 얻어 총 8개 영역의 세포를 분석했다.
각 세포의 분포와 역할을 파악하기 위해 ‘코덱스(CODEX, co-detection by indexing)란 분석 기법을 사용됐다. 특정한 단백질과 결합해 조직을 반복적으로 염색하는 물질을 사용해 각 세포 유형을 확인하는 방식이다. 연구팀은 코덱스 분석 기법을 통해 식별된 세포를 유형별로 나눈 뒤 리보핵산(RNA)를 포함한 염색체 정보를 분석했다.
분석한 결과 장 내벽을 구성하는 다양한 세포의 역할과 위치가 확인됐다. 상피세포, 결합조직세포, 신경세포, 면역세포 등이 특정한 질병이 생겼을 때 어떻게 운용되는지 규명했다.
산자이 자인 미국 워싱턴대 의대 교수 연구팀은 건강한 사람 45명의 신장과 질환을 앓고 있는 48명의 신장의 세포를 분석했다. 대부분의 질환은 노폐물을 배설하고 체내항상성을 유지하는 기관인 신장의 손상으로 이어진다. 신장 기능에 영향을 미치는 세포의 역할을 아는 것이 중요한 이유다.
연구팀은 신장에 존재하는 주요 세포 51종의 분포와 역할을 확인했다. 연구팀은 신장이 손상됐을 때 어떤 세포에 결함이 생기고 어떻게 복구되는지 확인한 점을 이번 연구의 주된 성과로 꼽았다.
마이클 앤절로 미국 스탠퍼드대 교수 연구팀은 임신 초기 상태 태반에 존재하는 세포들의 지도를 만들었다. 태반은 태아와 모체의 자궁벽을 연결해 영양 공급, 가스교환, 노폐물 배출 등의 기능을 담당하는 기관이다.
연구팀은 태반에서 채취한 50만개의 조직 샘플에서 588개의 세포를 분석했다. 임신 6~20주 각 시기별 세포와 태반 간의 상호작용을 규명한 것이 주된 성과다. 각 연구를 이끈 연구팀은 이번에 확인된 세포 지도가 실질적인 인간 건강의 증진으로 이어지기 위해선 질병과 각 세포의 강력한 연관성을 확인하는 후속 연구가 필요하다고 말했다.