
심해의 열수분출공 주변은 미생물이 살아갈 수 있는 열과 영양분을 공급한다. 최근 일본 연구자들이 수심 2533미터 해저에서 채취한 토양 시료에 존재하는 아스가드 고세균을 실험실에서 배양하는 데 성공해 화제가 되고 있다. ‘네이처’ 제공
얼마 전 국제학술지 ‘네이처’를 뒤적이던 필자는 이달 15일자 기사를 보고 눈이 번뜩 뜨였다. 일본 과학자들이 무려 12년에 걸친 각고의 노력 끝에 아스가드(Asgard) 고세균을 실험실에서 배양하는 데 성공했다는 내용이다.
미생물 한 종을 배양한 게 뭐 그리 대단한 성과냐고 의아해할 독자도 있겠지만 이게 보통 미생물이 아니다. 어쩌면 생명과학의 미스터리인 진핵생물의 기원을 설명할 수 있는 열쇠를 쥐고 있을지도 모르기 때문이다.
사실 논문은 아직 학술지에 실리지도 않았다. 지난 8월 6일 학술지에 투고하며(어딘지는 알려지지 않았다) 생명과학 분야의 논문 저장소인 ‘바이오 아카이브(bioRxiv)’에 올려 공개된 것이다. ‘네이처’가 투고 중인 논문을 뉴스로 다루는 건 드문 일이다. 그만큼 엄청난 연구결과라는 말이다.
필자는 귀가하자마자 저자들의 논문을 다운로드받아 읽어봤다. 실험 내용도 대단했지만 그 결과가 함축하는 바도 심오했다. 필자가 느낀 감동을 독자들도 공감하길 기대하며 아스가드 고세균 배양 성공의 의미에 대해 설명한다.
허 찔린 스웨덴 연구팀
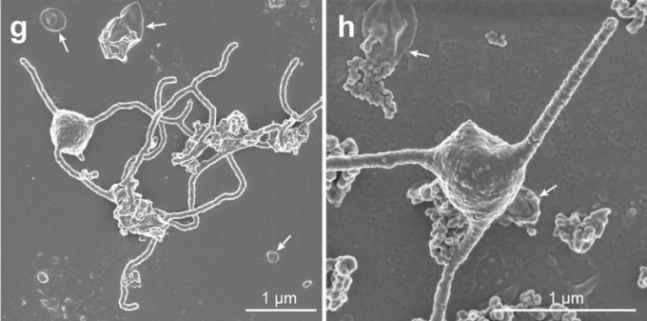
전자현미경으로 본 아스가드 고세균 MK-D1의 모습이다. 세포는 대장균보다도 작지만 실 같은 돌출물과 작은 주머니 형태인 소포(화살표)가 존재하는 특이한 형태다. ‘bioRxiv’ 제공
오늘날 생명과학의 최대 미스터리는 생물의 기원, 어떻게 무생물에서 생물이 나왔느냐 하는 것이다. 그다음 미스터리가 바로 원핵생물에서 어떻게 진핵생물이 진화했느냐 아닐까. 그 뒤 단세포 진핵생물에서 다세포 진핵생물이 진화했고 궁극적으로 사람이 등장했기 때문이다.
진핵생물의 기원을 설명하는 가설의 전개과정을 다 되짚어보려면 이번 연구를 다루기도 전에 지칠 것이므로 생략하고 오늘날 많은 사람들이 인정하는 시나리오만 소개한다. 과거 어느 시점에서(대략 20억 년 전으로 추정) 원핵생물인 고세균(아케아)이 또 다른 원핵생물인 진정세균(박테리아)를 포획했다. 포획된 박테리아는 세포에 에너지를 공급하는 세포소기관인 미토콘드리아가 됐고 고세균의 게놈이 막에 둘러싸이면서 진핵세포로 진화했다는 것이다.
이 시나리오는 고세균의 게놈이 같은 원핵생물인 박테리아의 게놈보다 진핵생물의 게놈과 더 가깝다는 발견에 기반한다. 그럼에도 그 차이가 꽤 커서 20억 년 전 박테리아를 포획했던 고세균의 모습을 떠올리기는 어려웠다.
그런데 2015년 ‘네이처’에 놀라운 연구결과가 실렸다. 스웨덴 웁살라대 티스 에타마 교수팀이 수심 3283미터에 이르는 북극해의 해저 토양을 채취해 메타게놈을 분석한 결과 기존 고세균과 상당히 다르고 오히려 진핵생물과 더 가까운 새로운 고세균의 존재를 확인했다는 것이다.
메타게놈(metagenome)이란 시료에 있는 생물체 전체의 게놈이다. 예전에는 실험실에서 배양하는 데 성공한 미생물만 게놈을 분석할 수 있었지만, 감도가 높은 분석법이 나오고 생물정보학이 발전하면서 시료 상태에서 게놈을 밝힐 수 있다. 연구자들은 이 고세균을 위해 ‘로키아케오타(Lokiarchaeota)’라는 새로운 문(phylum)을 만들었다.
‘로키’로 불리는 이 고세균의 게놈에는 지금까지 진핵생물에만 있는 것으로 알려진 유전자가 175개나 존재했다. 이어지는 메타게놈 연구에서 여러 종이 추가로 발견돼 로키 외에도 네 가지 문이 더 만들어졌다. 게놈 비교분석 결과 이 가운데 헤임달라케오타(Heimdallarchaeota)가 진핵생물과 가장 가까운 것으로 나타탔다. 연구자들은 2017년 역시 ‘네이처’에 발표한 논문에서 이들을 포함하는 아스가드 상문(Asgard superphylum)을 제안했다.
따라서 약 20억 년 전 아스가드 상문에 속하는 고세균(아마도 헤임달라케오타)이 박테리아를 포획해 진핵생물로 향하는 진화의 여정을 시작했을 가능성이 높다. 그럼에도 심해처럼 극한의 환경에서 자라는 미생물은 대부분 실험실에서 배양이 되지 않기 때문에 아스가드 고세균이 어떻게 생겼는지조차 알지 못하는 상태였다.
그런데 이번에 일본해양지구과학기술연구소를 비롯한 일본 공동연구자들이 아스가드 고세균을 배양하는 데 성공했다고 발표한 것이다. 논문에는 물론 고세균의 전자현미경 사진도 실려있다. 아스가드 고세균 연구를 이끌었고 십중팔구 배양을 시도하고 있었을 스웨덴 웁살라대 연구팀으로서는 불의의 일격일 것이다. 그런데 어떻게 일본 연구자들은 아스가드 고세균의 존재가 알려지지도 않은 12년 전에 배양실험을 시작한 것일까.
크기는 작지만 독특한 구조 지녀
사실 이들은 심해 해저 토양을 채취해 실험실에 비슷한 환경의 배양기를 만들어 배양을 시도한 것이다. 산소가 없는 조건에서 메탄을 공급해 이를 먹고 살아가는 미생물과 (혹시 있다면) 이들과 공생하는 미생물을 증식시켜 실체를 밝혀볼 계획이었다.
연구자들은 2006년 5월 6일 일본 남쪽 바다 난카이 해구의 수심 2533미터 해저에서 채취한 토양 시료를 배양기에 넣고 10℃에서 무려 2013일을 배양했다. 5년 반에 이르는 긴 기간이다. 해저 미생물은 증식이 워낙 더뎌 두 배로 늘어나는 데 수개월이 걸릴 수도 있기 때문이다. 반면 대장균 같은 미생물은 조건만 맞으면 20분 만에 두 배가 된다.
그 뒤 배양액을 시험관에 나눠 다시 여러 조건에서 1년 동안 배양했다. 배양액이 약간 뿌옇게 된(증식했다는 뜻이다) 시험관에서 시료를 채취해 메타게놈을 분석한 결과 아스가드 고세균을 비롯해 다양한 미생물이 존재했다. 다만 철저하게 분석하지 않았기 때문에 당시는 아스가드 고세균가 얼마나 중요한 미생물인지 몰랐다(물론 이름도 붙이지 않았다).
그 뒤 연구자들은 다양한 조건에서 배양을 계속했고 이 과정에서 스웨덴 연구팀이 발표한 아스가드 고세균을 자신들이 키우고 있다는 사실을 알아차렸을 것이다. 이 가운데 한 종이 증식이 잘 돼 실체를 규명할 수 있었고 연구자들은 ‘프로메테오아케움 신트로피쿰(Prometheoarchaeum syntrophicum)’이라는 학명을 붙인 뒤 균주 ‘MK-D1’으로 불렀다. 녀석의 온전한 게놈을 해독한 결과 ‘로키’에 속하는 것으로 밝혀졌다.
전자현미경으로 들여다본 MK-D1의 모습은 실망스러웠다. 오늘날 아스가드 고세균이 20억 년 전 (미토콘드리아가 될) 박테리아를 식작용(phagocytosis)으로 포획했을 고세균과 비슷하다면 덩치가 꽤 커야 하는 데 1㎛(마이크로미터. 1㎛는 100만분의 1미터)도 안 돼 대장균보다도 작았기 때문이다. 박테리아를 잡아먹기는커녕 잡아먹힐 크기다!
그러나 자세히 들여다보자 MK-D1의 생김새가 예사롭지 않았다. 세포 표면에 짧은 줄 같은 돌출물이 여럿 있었다. 얼핏 신경세포(뉴런)처럼 보인다. 또 표면 막에 소포(vesicle)가 존재했다. 나중에 연구자들이 제시하는 진핵생물 진화 시나리오에서 이 돌출물과 소포가 중요한 역할을 한다.
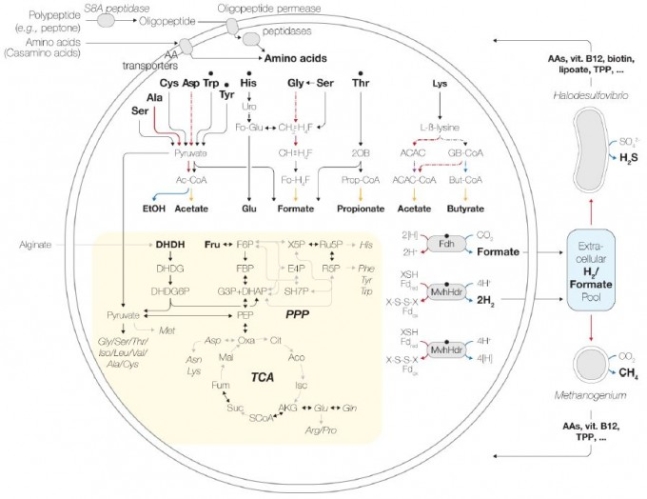
MK-D1의 배양 조건과 게놈의 대사 관련 유전자 분석을 토대로 제시한 대사 메커니즘과 미생물 두 종과의 공생영양 관계를 보여주는 도식이다. 왼쪽 큰 원이 MK-D1이고 오른쪽 위가 황산염 환원 박테리아, 오른쪽 아래가 메탄 생성 고세균이다. ‘bioRxiv’ 제공
공생 파트너는 박테리아와 고세균
MK-D1의 가장 흥미로운 특성 가운데 하나는 혼자서는 증식하지 못한다는 것이다. 파트너 미생물이 존재해야 증식할 수 있다. 이런 관계를 영양공생(syntrophism)이라고 부른다. 연구자들이 학명을 신트로피쿰이라고 붙인 이유다. MK-D1이 영양공생으로 살아간다는 건 매우 중요한 발견이다. 고세균이 공생하던 박테리아를 포획했다는 게 진핵생물 진화 시나리오의 핵심이기 때문이다.
여러 배양조건에서 실험한 결과 MK-D1은 아미노산을 먹이로 삼아 이를 분해해 에너지를 얻는 것으로 밝혀졌다. MK-D1 게놈을 해독해 대사 관련 유전자를 분석한 결과도 이 관찰을 뒷받침했다. 아미노산 분해 과정에서 나오는 전자는 수소분자나 포름산염 형태로 배출되고 가까이 있는 공생 미생물이 이를 이용해 살아나간다. 한편 MK-D1은 공생 미생물이 만든 아미노산과 비타민 등을 흡수해 살아나간다.
그런데 막상 공생하는 미생물의 실체를 보니 진핵세포 기원 시나리오에서 미토콘드리아가 된 것으로 추정하는 알파프로테오박테리움(alphaproteobacterium)이 아니었다. 대신 황산염을 환원시키는 박테리아와 메탄을 생성하는 고세균이었다. 추가 배양실험 결과 박테리아는 없어도 되지만 메탄 생성 고세균은 꼭 있어야 한다는 사실이 밝혀졌다.
고세균 두 종(MK-D1과 메탄 생성 고세균)이 영양공생 관계이고 따라서 이 자체로는 진핵세포 진화와 직접적인 관련이 없다. 그럼에도 연구자들은 배양과 게놈 데이터를 바탕으로 진핵세포 기원을 설명하는 가설인 ‘E3 모형’을 만들었다. E3은 Entangle(얽힘), Engulf(삼킴), Enslave(예속)를 뜻한다.
배양된 고세균 보고 새로운 시나리오 만들어
연구자들은 진핵생물의 진화를 여섯 단계로 나눠 제시했다. 첫 단계는 광합성을 하는 시아노박테리아의 등장으로 산소 농도가 높아지기 시작한 27억 년 전(추정) 이전에 일어났을 것이다. 오늘날 심해 해저 토양의 조건과 비슷한, 산소가 희박하고 아미노산 같은 유기물이 존재하는 원시 지구의 환경에서 아스가드 고세균은 메탄 생성 고세균과 영양을 주고받으며 근근이 살아갔을 것이다. 오늘날 MK-D1의 생태에서 당시 모습을 엿볼 수 있다는 말이다.
27억 년 전부터 광합성으로 산소의 농도가 올라가기 시작하면서 아스가드 고세균 가운데 산소가 존재에 적응하는 종류가 생겨났다. 바로 헤임달라케오타(줄여서 헤임달)다. 이 환경에서는 황산염을 비롯한 유기물이 풍부했으므로 황산염환원박테리아가 주된 공생 파트너였을 것이고 아울러 산소를 이용해(산화반응) 에너지를 만드는 알파프로테오박테리움과 공생도 시작했을 것이다. 헤임달 역시 박테리아를 잡아먹기에는 너무 작았을 것이므로 표면에 난 긴 돌기로 감싸 가까이 뒀을 것이다.
광합성이 본격화되면서 산소 농도가 점점 높아지자 알파프로테오박테리움이 점점 더 중요해졌다. 아스가드 고세균 표면의 돌출물과 소포가 박테리아와 얽히면서(Entangle) 서로 뗄 수 없는 사이가 되고 결국 삼키게 되는 게(Engulf) 세 번째 단계다.
박테리아를 삼킨 고세균에서 핵막이 형성되는 게 네 번째 단계이고 박테리아 게놈 대부분이 고세균 게놈으로 넘어가면서 삼켜진 박테리아가 독립성을 잃어가는 과정이 다섯 번째 단계다. 이제 박테리아는 완전히 예속돼(Enslave) 세포소기관 미토콘드리아가 돼 세포가 흡수한 영양분을 산화시켜 에너지 분자(ATP)를 만드는 일을 전담하는 게 마지막 단계다.
이번 ‘로키’ 아스가드 고세균 배양 성공으로 알게 된 지식을 바탕으로 제시된 E3 모형이 과연 맞는 것인지는 아직 알 수 없지만 필자 눈에는 꽤 그럴듯하게 보인다. 차후에 ‘헤임달’ 고세균도 배양에 성공한다면 E3 모형의 두 번째와 세 번째 단계를 실험으로 확인해 볼 수도 있지 않을까. 지금까지 이론 생물학의 영역에 머물렀던 진핵세포 진화 연구가 실험 생물학으로 넘어가고 있다.
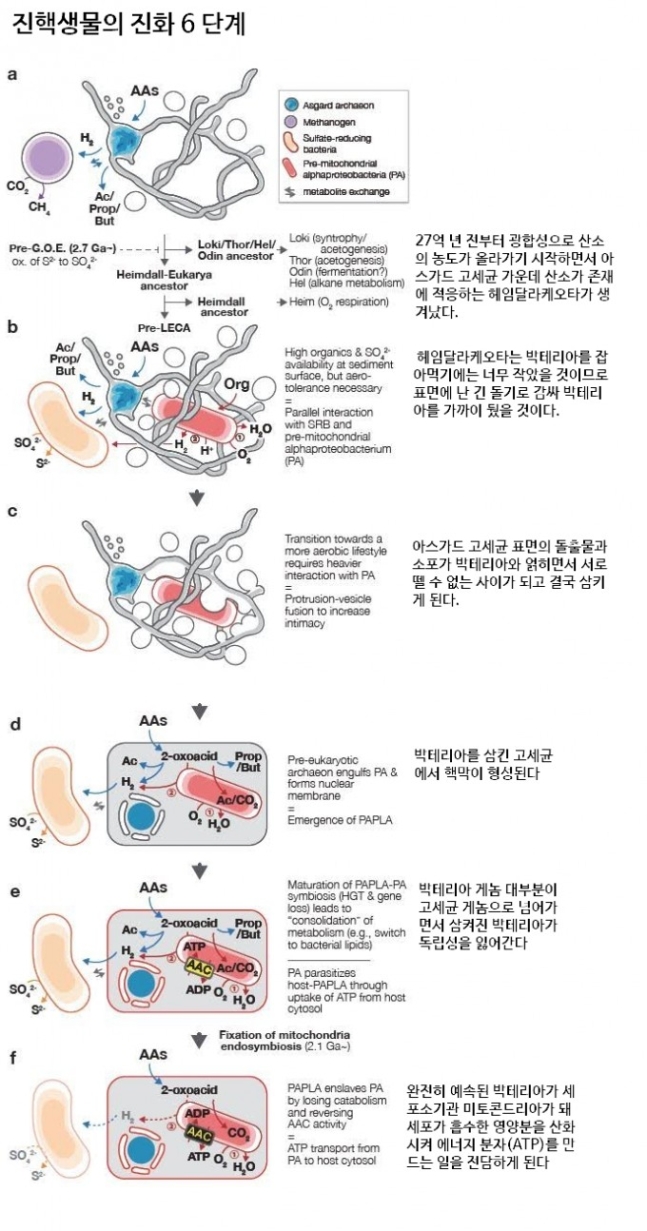
연구자들은 이번 MK-D1의 발견을 토대로 진핵생물의 기원을 설명하는 E3 모형을 만들었다. 이를 여섯 단계로 설명하는 도식으로 자세한 설명은 위 본문을 참조하라. ‘bioRxiv’ 제공
(원문: 여기를 클릭하세요~)