A tenet of elementary biology is that mitochondria — the cell’s powerhouses — and their DNA are inherited exclusively from mothers. A provocative study suggests that fathers also occasionally contribute.
The DNA of eukaryotic organisms (such as animals, plants and fungi) is stored in two cellular compartments: in the nucleus and in organelles called mitochondria, which transform nutrients into energy to allow the cell to function. The nucleus harbours most of our genes, tightly packaged into 46 chromosomes, of which half are inherited from our mother’s egg and half from our father’s sperm. By contrast, mitochondrial DNA (mtDNA) was thought to derive exclusively from maternal egg cells, with no paternal contribution1. Writing in Proceedings of the National Academy of Sciences, Luo et al.2 challenge the dogma of strict maternal mtDNA inheritance in humans, and provide compelling evidence that, in rare cases, the father might pass on his mtDNA to the offspring, after all.
Human eggs contain more than 100,000 copies of mtDNA, whereas sperm contain approximately 100 copies3. Early hypotheses suggested that paternal mtDNA molecules became diluted in number relative to maternal mtDNA ones in the fertilized egg, but these ideas were replaced when evidence from various organisms, such as the uni-cellular alga Chlamydomonas reinhardtii4 and medaka fish5, showed that paternal mtDNA is rapidly eliminated after fertilization. For decades, researchers have speculated on why healthy organisms obtain their cellular powerhouses from just one parent and on the possible evolutionary advantages conferred by mitochondrial genes inherited in this fashion.
A healthy individual’s mtDNA molecules are mostly identical. But in people with diseases caused by mtDNA mutations, normal and mutant mtDNA molecules typically coexist in a single cell — a situation termed heteroplasmy6. Disease severity is often associated with the amount of mutant mtDNA in cells, which is in turn determined by events that occurred when the person’s mother was still an embryo7. The developing eggs in the female embryo go through an ‘mtDNA bottle-neck’, in which the number of mtDNA copies is first reduced and then amplified to more than 100,000 copies8,9. Accordingly, variable amounts of mutant and normal mtDNA are present in the mature eggs of an individual woman, and, therefore, in the cells of her offspring. This phenomenon influences the severity of diseases caused by mtDNA mutations, and can lead to very different manifestations between individuals from the same family7.
Luo and colleagues identified three families with mtDNA heteroplasmy that could not be explained by maternal inheritance. The story started with a young boy suspected of having a mitochondrial disease. The authors performed high-resolution mtDNA sequencing, but did not identify any disease-causing mtDNA mutations. However, their analysis uncovered unusually high levels of mtDNA heteroplasmy. Intriguingly, the same unusual pattern of mtDNA variation was found in the boy’s mother and in his two healthy sisters (Fig. 1).
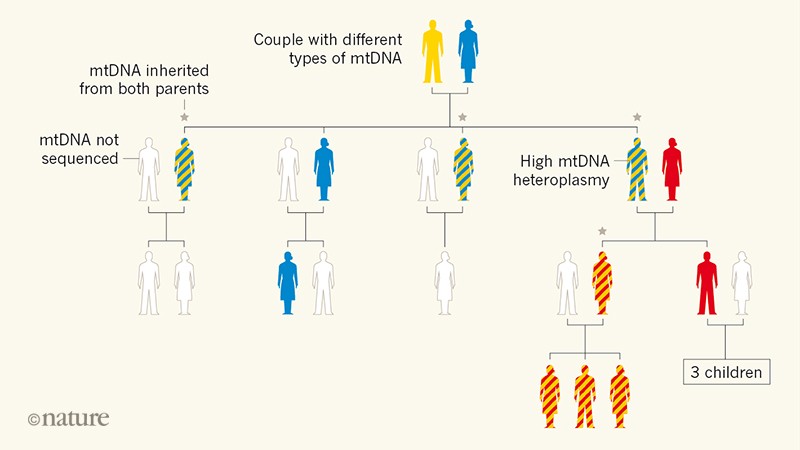
Figure 1 | Family tree revealing paternal inheritance of mitochondrial DNA (mtDNA). Luo et al.2sequenced the mtDNA of several members of a family in which many individuals had a high level of mtDNA heteroplasmy (the presence of distinct genetic variants in the same cell). This mtDNA variability is denoted by two colours in the same silhouette of an individual. The analysis showed that some of the individuals with heteroplasmy had inherited mtDNA from both of their parents, breaking the usual pattern of exclusive maternal inheritance of mtDNA. Luo et al. suggest that the ability to inherit paternal mtDNA is a genetic trait.
To trace the origin of this mysterious mtDNA pattern, Luo et al. extended their investigation to the previous generation. Sequencing of the mtDNA of the boy’s maternal grandparents revealed an unexpected contribution: his unusual mtDNA pattern seemed to be the product of mtDNA from both grandparents. The authors went on to identify two additional and unrelated families that had biparental mitochondrial transmission. A similar scenario was previously observed in an individual with mitochondrial disease who had a paternally inherited mtDNA variant10. Together, these reports provide evidence for biparental mitochondrial inheritance in humans.
Human disease-causing mtDNA mutations were originally reported in 1988 (refs 6, 11)6,11, and more than 200 such mutations (see go.nature.com/2fucdqt) have been discovered since then, most of them occurring in a hetero-plasmic context7. More-over, the estimated frequency of mutations of matrilineal mtDNA has made it a useful and often-used tool in studies of ancestry and evolution, as well as in forensic identification12. Human mtDNA has also been a valuable tool in archaeology, because its small size (16,569 base pairs) and circular form make it more resistant to degradation than is nuclear DNA (which has around 3 billion base pairs)13.
Given this long and multifaceted research history, why would paternal mtDNA have remained undetected? Luo et al. suggest that mtDNA heteroplasmy is often overlooked in diagnostics when it does not involve a disease-causing variant. Although this might be true to some extent, it is a rather unsatisfactory explanation in this era of deep DNA sequencing. Nevertheless, Luo and colleagues’ findings should provoke a re-assessment of the extensive global mtDNA sequencing data available, for those wishing to unearth further instances of atypical heteroplasmy. If the paternal contribution to mtDNA is more common than previously realized, this could alter some estimated timings of human evolution, because these are often based on predictions of mtDNA sequence variation under the assumption of exclusive maternal inheritance.
Although biparental inheritance of mtDNA and heteroplasmy coincided with disease symptoms in some of the individuals studied by Luo et al., the authors’ data do not demonstrate a causal link with disease. In fact, we cannot be certain that the study participants have mitochondrial disease, because no specific examinations to confirm this diagnosis are reported. Further study is needed to identify more cases of potential paternal mtDNA inheritance, and to determine the functional consequences of such heteroplasmy. Notably, this knowledge is relevant to mitochondrial-donation therapy (“three-parent babies”), which aims to prevent the transmission of disease-causing mtDNA to offspring14, but which can also potentially generate individuals with two types of mtDNA, one from the donor and another from the mother.
Could the amount of paternal mtDNA in a fertilized egg or developing embryo be deliberately boosted to diminish the adverse effects of mutant maternal mtDNA when this is present? This is an interesting option, but still far from reality. In addition to evading elimination, paternal mtDNA molecules would need to have a considerable replicative advantage over maternal ones to reach meaningful proportions.
Will Luo and colleagues’ findings affect the counselling of individuals carrying disease-causing mtDNA mutations who are considering having children? Not greatly, because paternal mitochondrial transmission seems to be exceedingly rare in humans. At present, this discovery represents an interesting conceptual breakthrough, rather than one that will directly influence clinical practice.
Previous work15 has shown that mitophagy, the process by which cells ‘eat’ their own mitochondria, has a role in the selective elimination of paternal mitochondria. Given our rapidly expanding knowledge of mammalian mitophagy in vivo16, these rare instances of paternal mtDNA transmission might be attributed to defective mitochondrial turnover. The inheritance pattern of paternal mtDNA in Luo and colleagues’ study suggests that a yet unidentified gene on one of the autosomes (non-sex chromosomes) is involved in eliminating paternal mitochondria. The families in whom paternal mtDNA inheritance was observed provide an exciting opportunity to decipher the signalling pathways that modulate paternal mitochondrial elimination and prevent biparental mitochondrial transfer.
Nature 565, 296-297 (2019)
(원문: 여기를 클릭하세요~)